

Transcriptomic profiling of tissue environments critical for post-embryonic patterning and morphogenesis of zebrafish skin
- PMID: 37695017
- PMCID: PMC10495112
- DOI: 10.7554/eLife.86670
Transcriptomic profiling of tissue environments critical for post-embryonic patterning and morphogenesis of zebrafish skin
Abstract
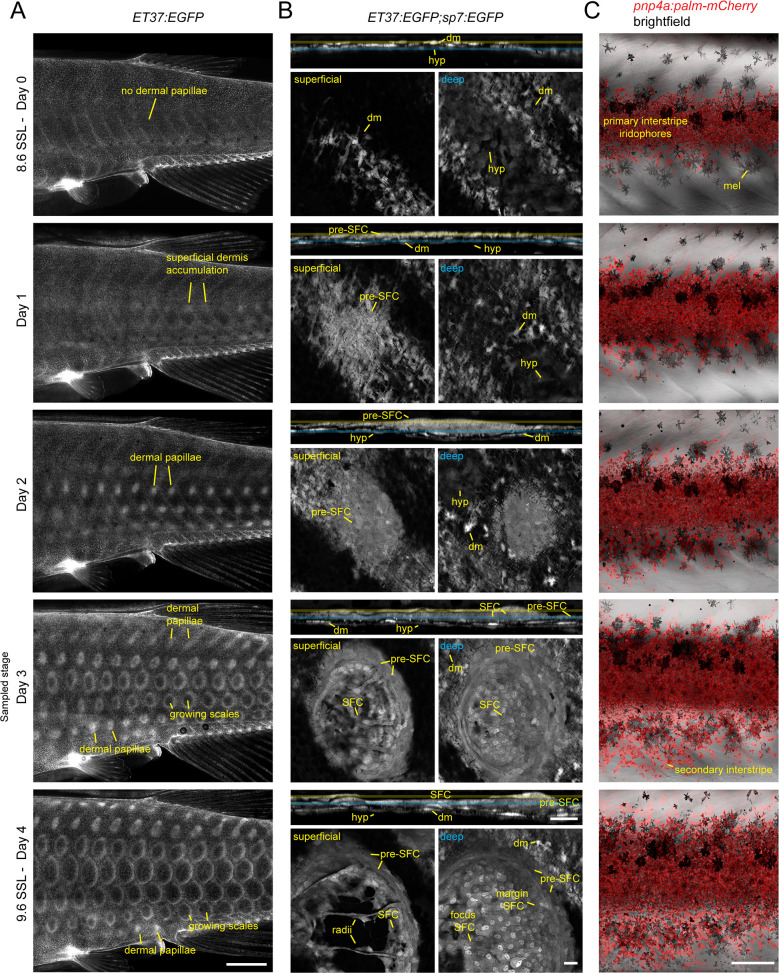
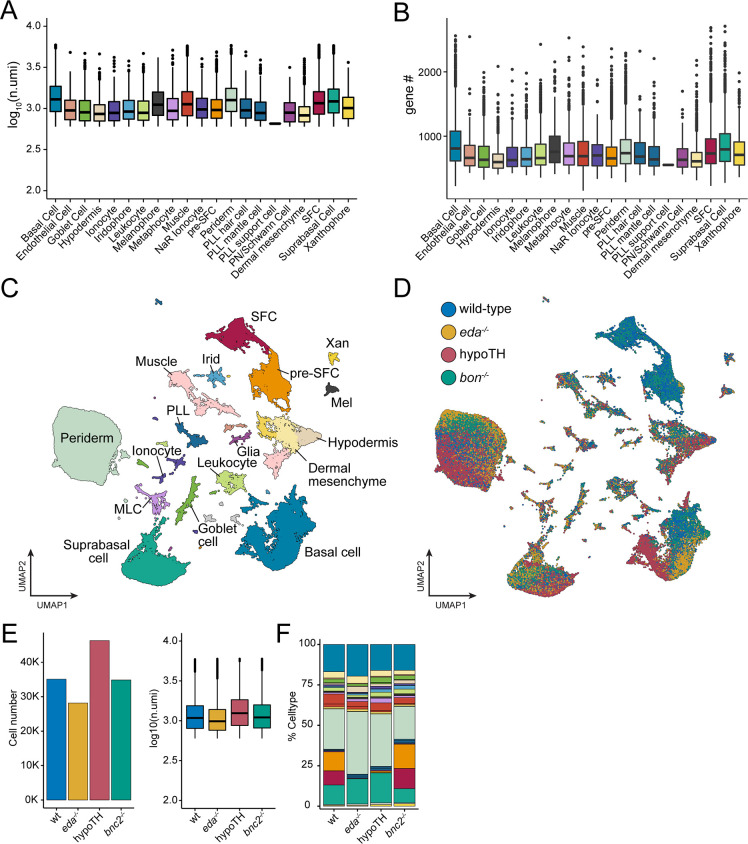
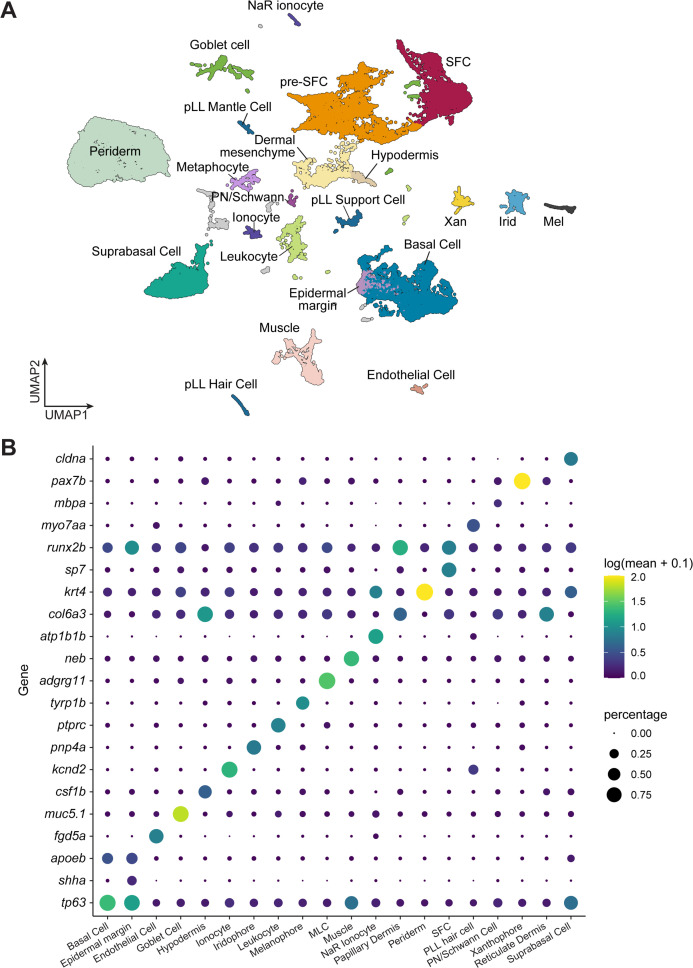
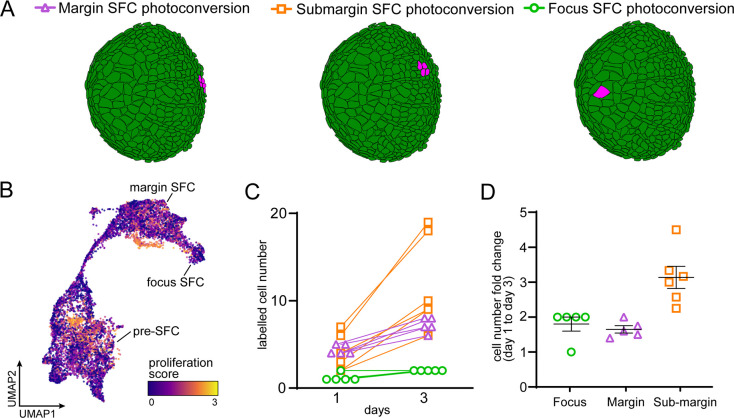
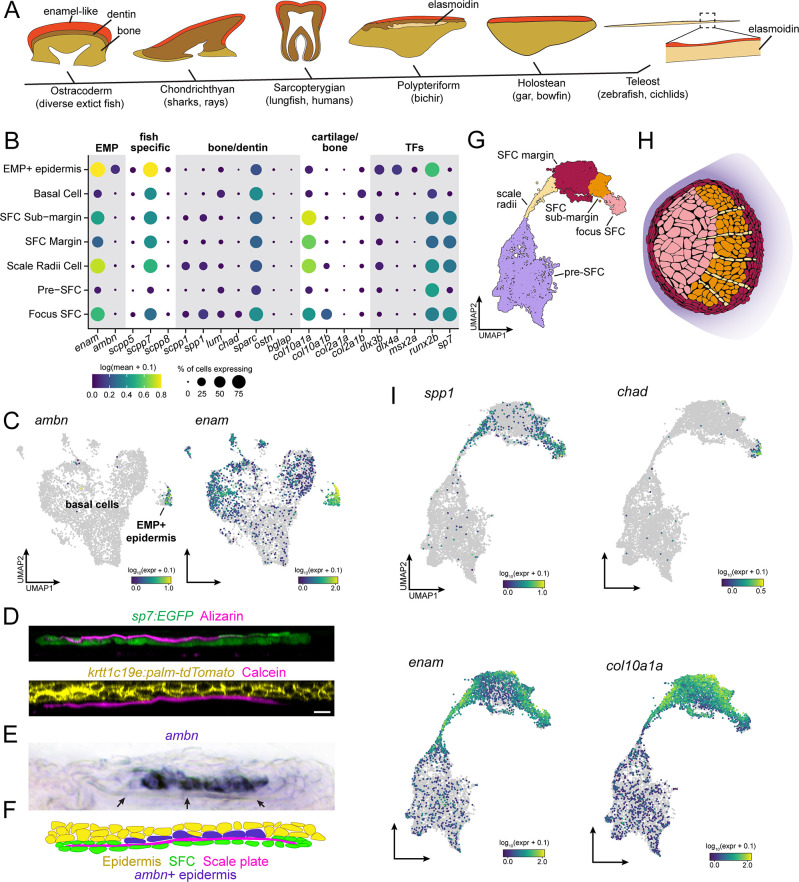
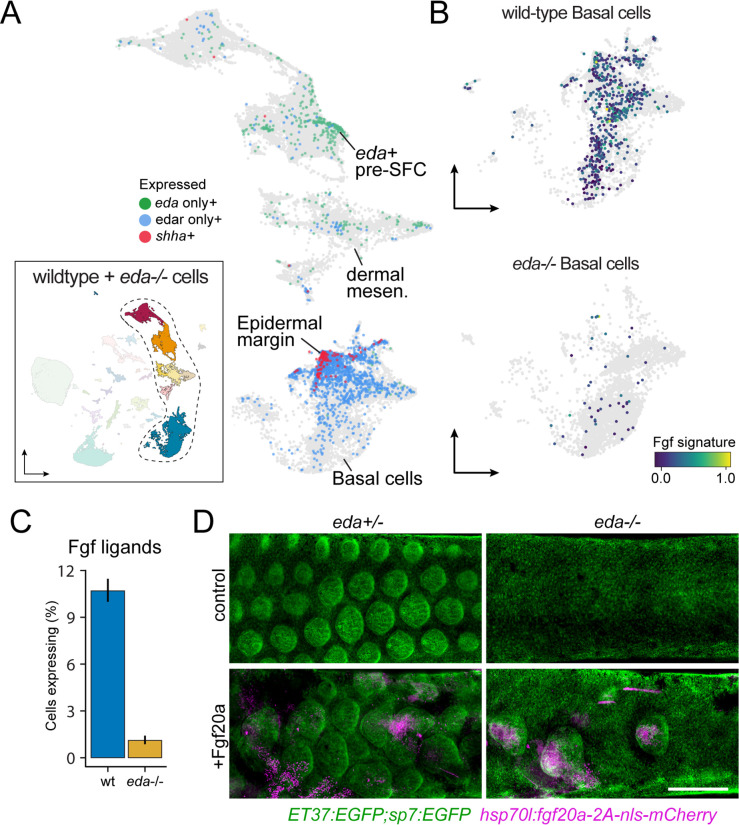
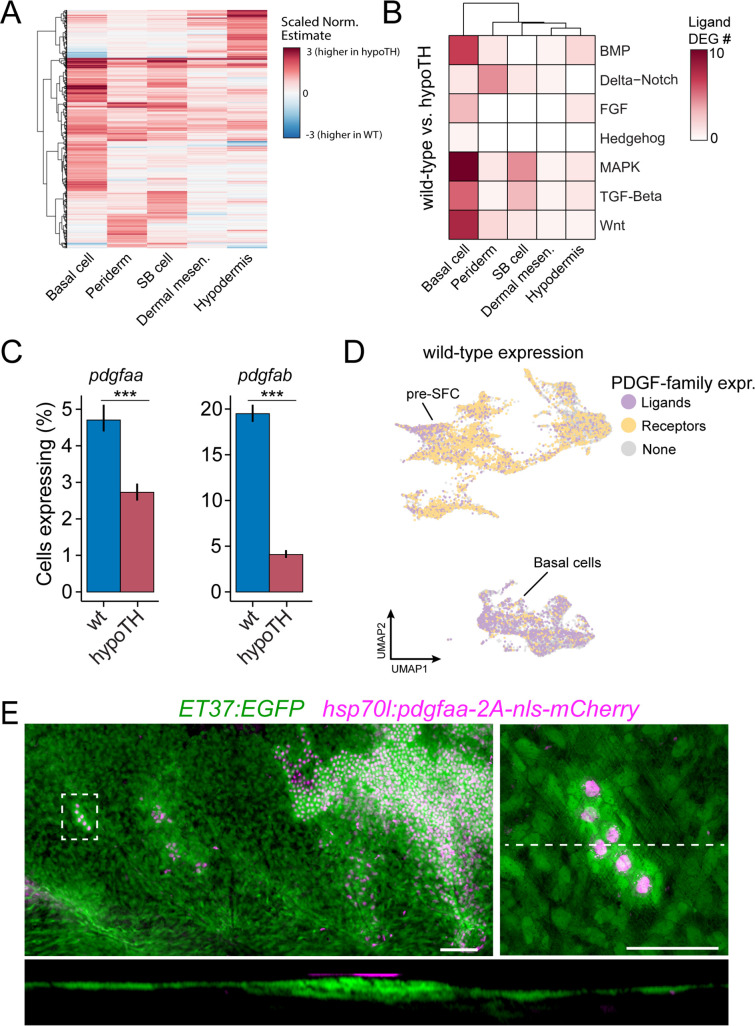
Pigment patterns and skin appendages are prominent features of vertebrate skin. In zebrafish, regularly patterned pigment stripes and an array of calcified scales form simultaneously in the skin during post-embryonic development. Understanding the mechanisms that regulate stripe patterning and scale morphogenesis may lead to the discovery of fundamental mechanisms that govern the development of animal form. To learn about cell types and signaling interactions that govern skin patterning and morphogenesis, we generated and analyzed single-cell transcriptomes of skin from wild-type fish as well as fish having genetic or transgenically induced defects in squamation or pigmentation. These data reveal a previously undescribed population of epidermal cells that express transcripts encoding enamel matrix proteins, suggest hormonal control of epithelial-mesenchymal signaling, clarify the signaling network that governs scale papillae development, and identify a critical role for the hypodermis in supporting pigment cell development. Additionally, these comprehensive single-cell transcriptomic data representing skin phenotypes of biomedical relevance should provide a useful resource for accelerating the discovery of mechanisms that govern skin development and homeostasis.
Keywords: adult form; developmental biology; differentiation; morphogenesis; pigment; skin; squamation; zebrafish.
Conflict of interest statement
AA, LS, AC, SS, CE, BC, DW, WP, CT, DP No competing interests declared
Figures










Update of
- doi: 10.1101/2021.05.12.443782
- doi: 10.7554/eLife.86670.1
- doi: 10.7554/eLife.86670.2
- doi: 10.7554/eLife.86670.3
Similar articles
-
The Developmental Genetics of Vertebrate Color Pattern Formation: Lessons from Zebrafish.Curr Top Dev Biol. 2016;117:141-69. doi: 10.1016/bs.ctdb.2015.12.012. Epub 2016 Feb 23. Curr Top Dev Biol. 2016. PMID: 26969976 Review.
-
Thyroid hormone regulates abrupt skin morphogenesis during zebrafish postembryonic development.Dev Biol. 2021 Sep;477:205-218. doi: 10.1016/j.ydbio.2021.05.019. Epub 2021 Jun 3. Dev Biol. 2021. PMID: 34089732 Free PMC article.
-
Scale development in fish: a review, with description of sonic hedgehog (shh) expression in the zebrafish (Danio rerio).Int J Dev Biol. 2004;48(2-3):233-47. doi: 10.1387/ijdb.15272389. Int J Dev Biol. 2004. PMID: 15272389 Review.
-
Galanin Signaling in the Brain Regulates Color Pattern Formation in Zebrafish.Curr Biol. 2020 Jan 20;30(2):298-303.e3. doi: 10.1016/j.cub.2019.11.033. Epub 2020 Jan 2. Curr Biol. 2020. PMID: 31902721 Free PMC article.
-
Wnt/β-catenin regulates an ancient signaling network during zebrafish scale development.Elife. 2018 Jul 17;7:e37001. doi: 10.7554/eLife.37001. Elife. 2018. PMID: 30014845 Free PMC article.
Cited by
-
Melanocyte lineage dynamics in development, growth and disease.Development. 2024 Aug 1;151(15):dev201266. doi: 10.1242/dev.201266. Epub 2024 Aug 2. Development. 2024. PMID: 39092608 Free PMC article. Review.
-
col1a2+ fibroblasts/muscle progenitors finetune xanthophore countershading by differentially expressing csf1a/1b in embryonic zebrafish.Sci Adv. 2024 Apr 5;10(14):eadj9637. doi: 10.1126/sciadv.adj9637. Epub 2024 Apr 5. Sci Adv. 2024. PMID: 38578990 Free PMC article.
-
Anatomy, development and regeneration of zebrafish elasmoid scales.Dev Biol. 2024 Jun;510:1-7. doi: 10.1016/j.ydbio.2024.03.001. Epub 2024 Mar 6. Dev Biol. 2024. PMID: 38458375
-
Revisiting ameloblastin; addressing the EMT-ECM axis above and beyond oral biology.Front Cell Dev Biol. 2023 Nov 13;11:1251540. doi: 10.3389/fcell.2023.1251540. eCollection 2023. Front Cell Dev Biol. 2023. PMID: 38020879 Free PMC article.
-
The hidden depths of zebrafish skin.Elife. 2023 May 23;12:e88597. doi: 10.7554/eLife.88597. Elife. 2023. PMID: 37218526 Free PMC article.
References
-
- Aman AJ, Parichy DM. In: The Zebrafish in Biomedical Research. Guillemin KJ, Kent ML, editors. Academic Press; 2020. Chapter 8 - Zebrafish Integumentary system; pp. 91–96. - DOI
-
- Ayoola AO, Zhang BL, Meisel RP, Nneji LM, Shao Y, Morenikeji OB, Adeola AC, Ng’ang’a SI, Ogunjemite BG, Okeyoyin AO, Roos C, Wu DD. Population genomics reveals incipient speciation, introgression, and adaptation in the african mona monkey (cercopithecus mona) Molecular Biology and Evolution. 2021;38:876–890. doi: 10.1093/molbev/msaa248. - DOI - PMC - PubMed
Publication types
MeSH terms
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
