

Comprehensive Dual- and Triple-Feature Intersectional Single-Vector Delivery of Diverse Functional Payloads to Cells of Behaving Mammals
- PMID: 32574559
- PMCID: PMC7687746
- DOI: 10.1016/j.neuron.2020.06.003
Comprehensive Dual- and Triple-Feature Intersectional Single-Vector Delivery of Diverse Functional Payloads to Cells of Behaving Mammals
Abstract
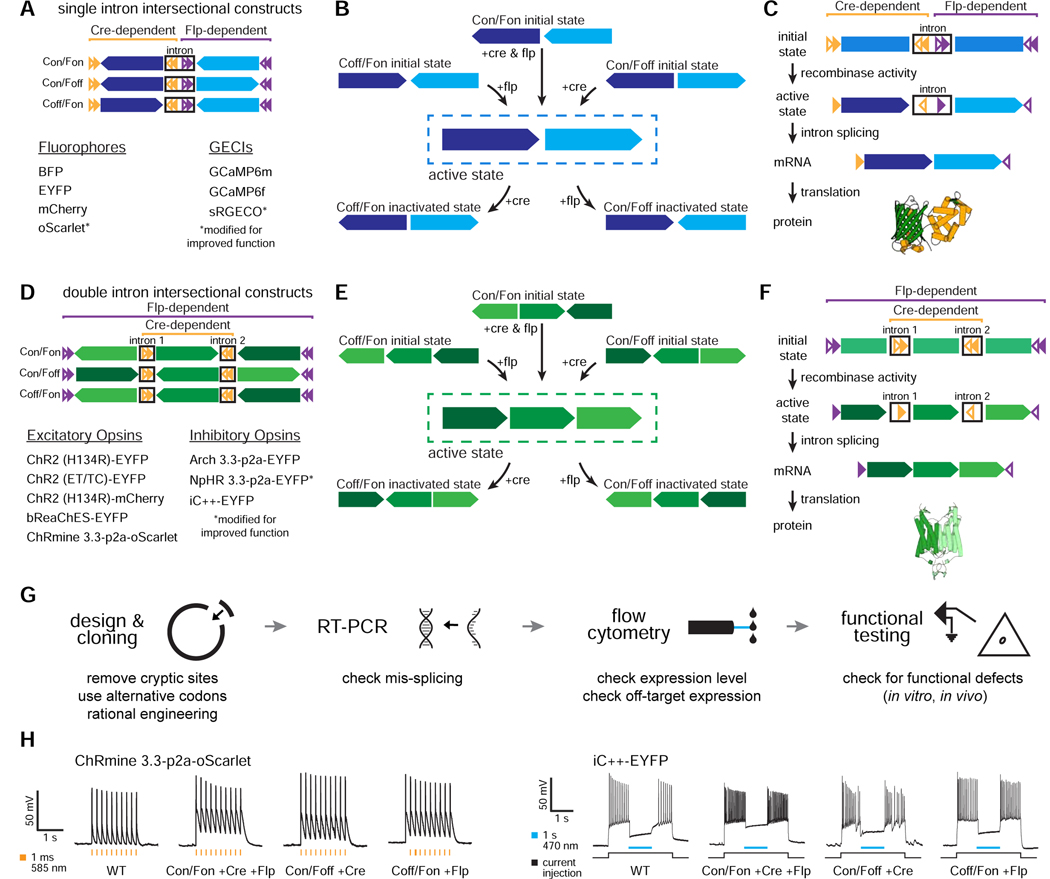
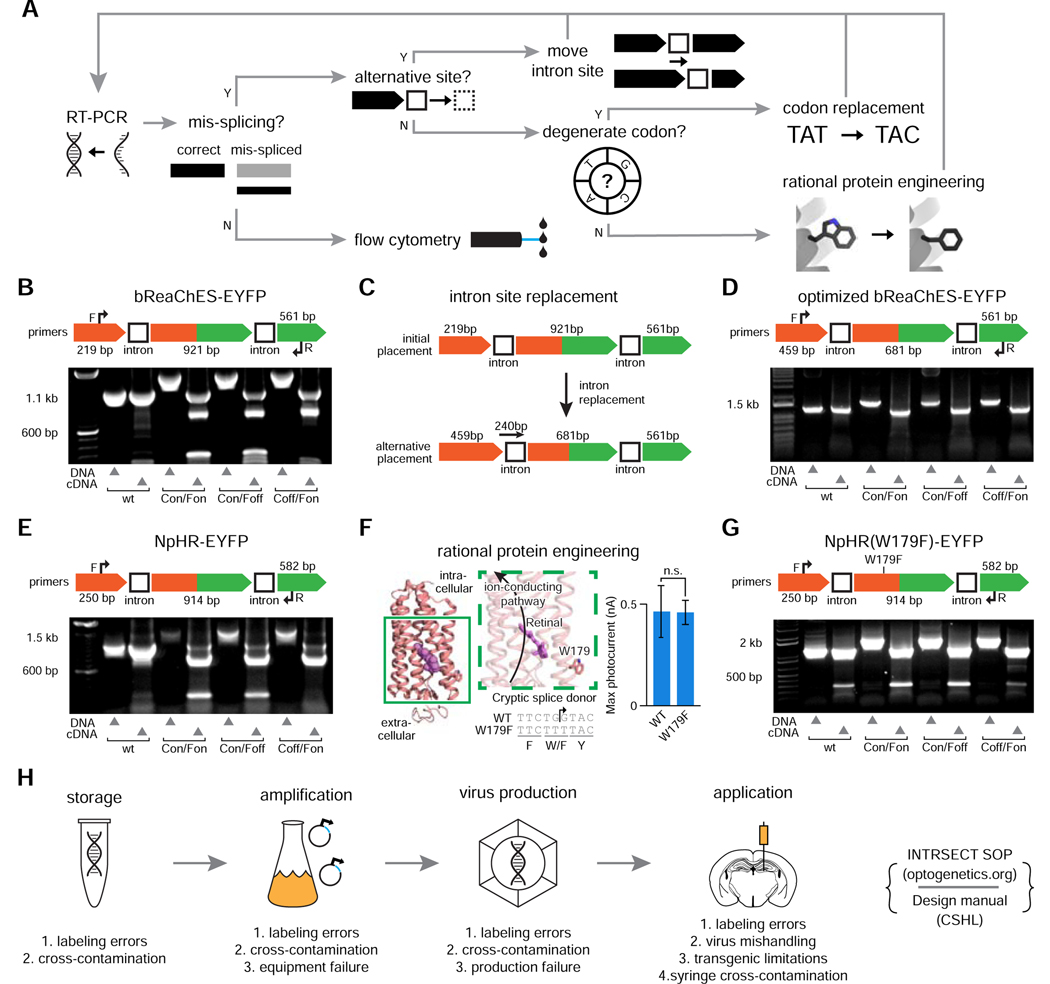
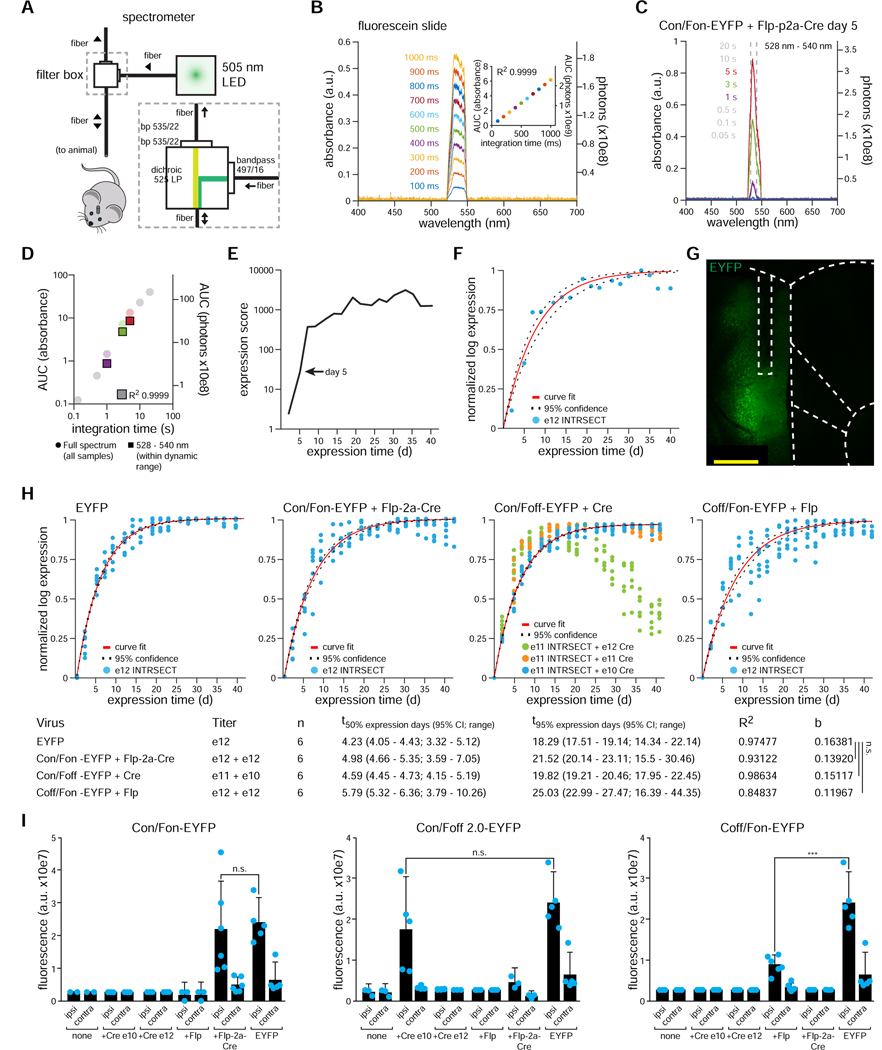
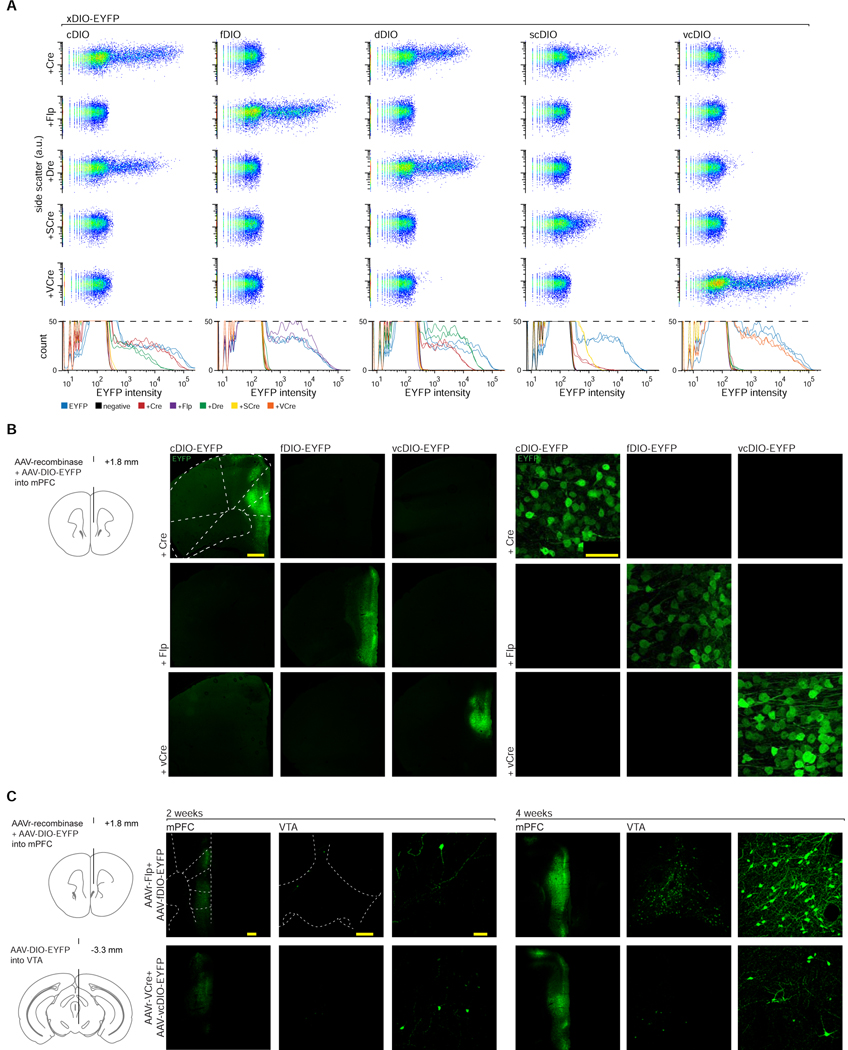
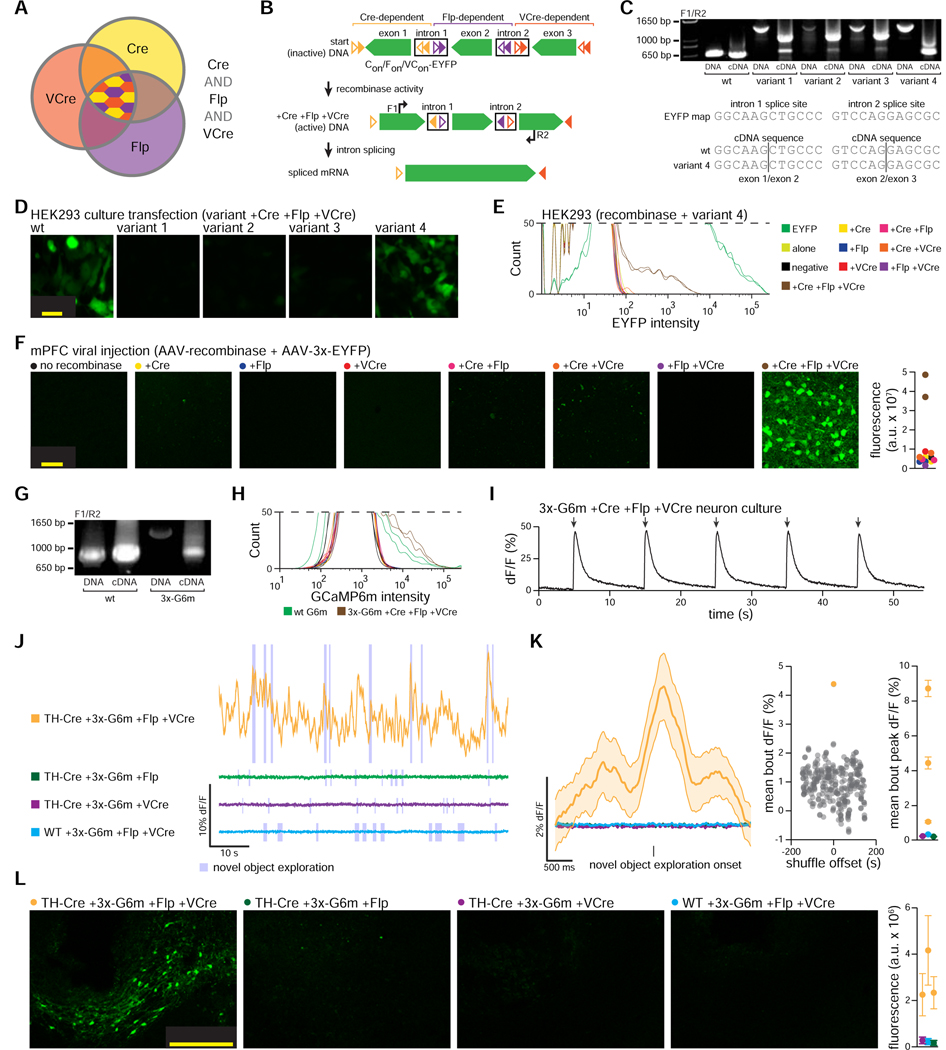
The resolution and dimensionality with which biologists can characterize cell types have expanded dramatically in recent years, and intersectional consideration of such features (e.g., multiple gene expression and anatomical parameters) is increasingly understood to be essential. At the same time, genetically targeted technology for writing in and reading out activity patterns for cells in living organisms has enabled causal investigation in physiology and behavior; however, cell-type-specific delivery of these tools (including microbial opsins for optogenetics and genetically encoded Ca2+ indicators) has thus far fallen short of versatile targeting to cells jointly defined by many individually selected features. Here, we develop a comprehensive intersectional targeting toolbox including 39 novel vectors for joint-feature-targeted delivery of 13 molecular payloads (including opsins, indicators, and fluorophores), systematic approaches for development and optimization of new intersectional tools, hardware for in vivo monitoring of expression dynamics, and the first versatile single-virus tools (Triplesect) that enable targeting of triply defined cell types.
Keywords: Cre; Flp; VCre; calcium imaging; channelrhodopsin; fluorophores; intersectional; optogenetics; recombinase; targeting.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests The authors declare no competing interests.
Figures
Similar articles
-
tTARGIT AAVs mediate the sensitive and flexible manipulation of intersectional neuronal populations in mice.Elife. 2021 Mar 11;10:e66835. doi: 10.7554/eLife.66835. Elife. 2021. PMID: 33704065 Free PMC article.
-
A CRISPR toolbox for generating intersectional genetic mouse models for functional, molecular, and anatomical circuit mapping.BMC Biol. 2022 Jan 28;20(1):28. doi: 10.1186/s12915-022-01227-0. BMC Biol. 2022. PMID: 35086530 Free PMC article.
-
A single-vector intersectional AAV strategy for interrogating cellular diversity and brain function.Nat Neurosci. 2024 Jul;27(7):1400-1410. doi: 10.1038/s41593-024-01659-7. Epub 2024 May 27. Nat Neurosci. 2024. PMID: 38802592
-
Challenges for Therapeutic Applications of Opsin-Based Optogenetic Tools in Humans.Front Neural Circuits. 2020 Jul 15;14:41. doi: 10.3389/fncir.2020.00041. eCollection 2020. Front Neural Circuits. 2020. PMID: 32760252 Free PMC article. Review.
-
Optogenetics: Illuminating the Future of Hearing Restoration and Understanding Auditory Perception.Curr Gene Ther. 2024;24(3):208-216. doi: 10.2174/0115665232269742231213110937. Curr Gene Ther. 2024. PMID: 38676313 Review.
Cited by
-
A versatile viral toolkit for functional discovery in the nervous system.Cell Rep Methods. 2022 May 26;2(6):100225. doi: 10.1016/j.crmeth.2022.100225. eCollection 2022 Jun 20. Cell Rep Methods. 2022. PMID: 35784651 Free PMC article.
-
Brain-wide mapping of efferent projections of glutamatergic (Onecut3+ ) neurons in the lateral mouse hypothalamus.Acta Physiol (Oxf). 2023 Jul;238(3):e13973. doi: 10.1111/apha.13973. Epub 2023 Apr 25. Acta Physiol (Oxf). 2023. PMID: 37029761 Free PMC article.
-
Gray areas: Neuropeptide circuits linking the Edinger-Westphal and Dorsal Raphe nuclei in addiction.Neuropharmacology. 2021 Oct 15;198:108769. doi: 10.1016/j.neuropharm.2021.108769. Epub 2021 Sep 2. Neuropharmacology. 2021. PMID: 34481834 Free PMC article. Review.
-
Dynamic and heterogeneous neural ensembles contribute to a memory engram.Curr Opin Neurobiol. 2021 Apr;67:199-206. doi: 10.1016/j.conb.2020.11.017. Epub 2020 Dec 31. Curr Opin Neurobiol. 2021. PMID: 33388602 Free PMC article. Review.
-
Deep imaging in the brainstem reveals functional heterogeneity in V2a neurons controlling locomotion.Sci Adv. 2020 Dec 4;6(49):eabc6309. doi: 10.1126/sciadv.abc6309. Print 2020 Dec. Sci Adv. 2020. PMID: 33277252 Free PMC article.
References
-
- Andrews BJ, Proteau GA, Beatty LG, and Sadowski PD (1985). The FLP recombinase of the 2μ circle DNA of yeast: Interaction with its target sequences. Cell 40, 795–803. - PubMed
-
- Asp M, Giacomello S, Larsson L, Wu C, Fürth D, Qian X, Wärdell E, Custodio J, Reimegård J, Salmén F, et al. (2019). A Spatiotemporal Organ-Wide Gene Expression and Cell Atlas of the Developing Human Heart. Cell 179, 1647–1660.e19. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
