

The mir-35 Family Links Maternal Germline Sex to Embryonic Viability in Caenorhabditis elegans
- PMID: 30679246
- PMCID: PMC6404603
- DOI: 10.1534/g3.118.200863
The mir-35 Family Links Maternal Germline Sex to Embryonic Viability in Caenorhabditis elegans
Abstract
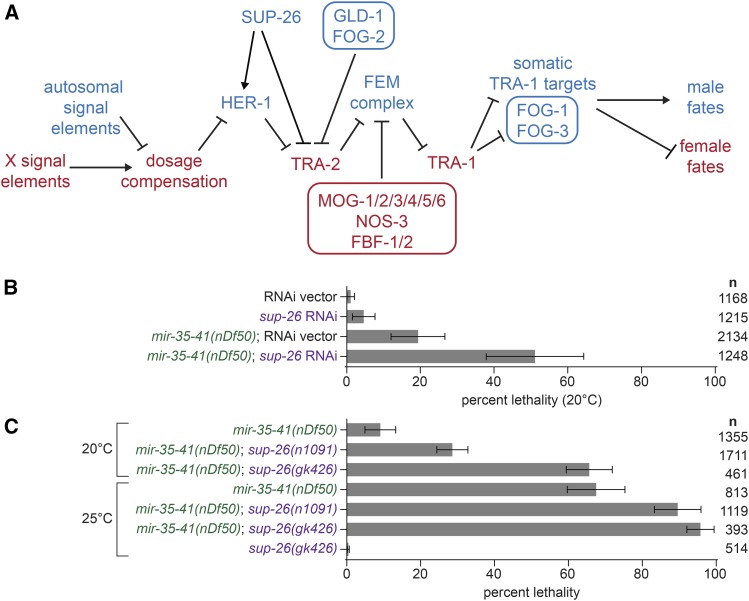
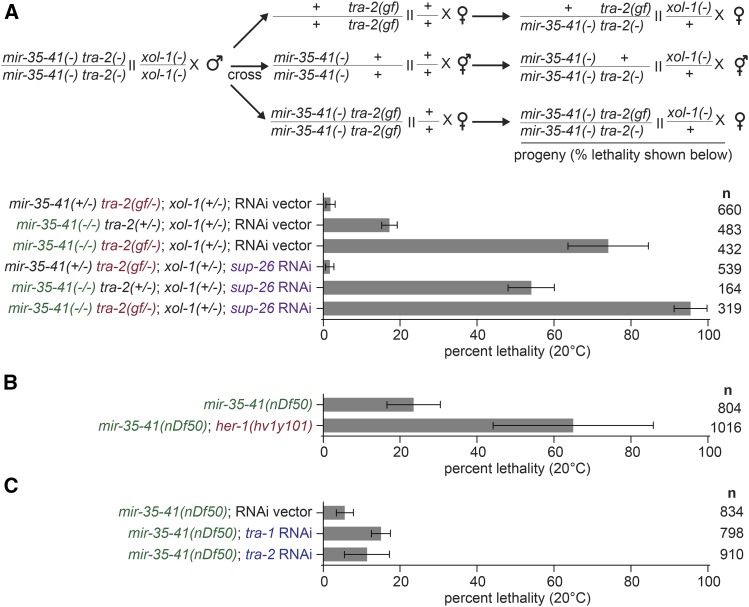
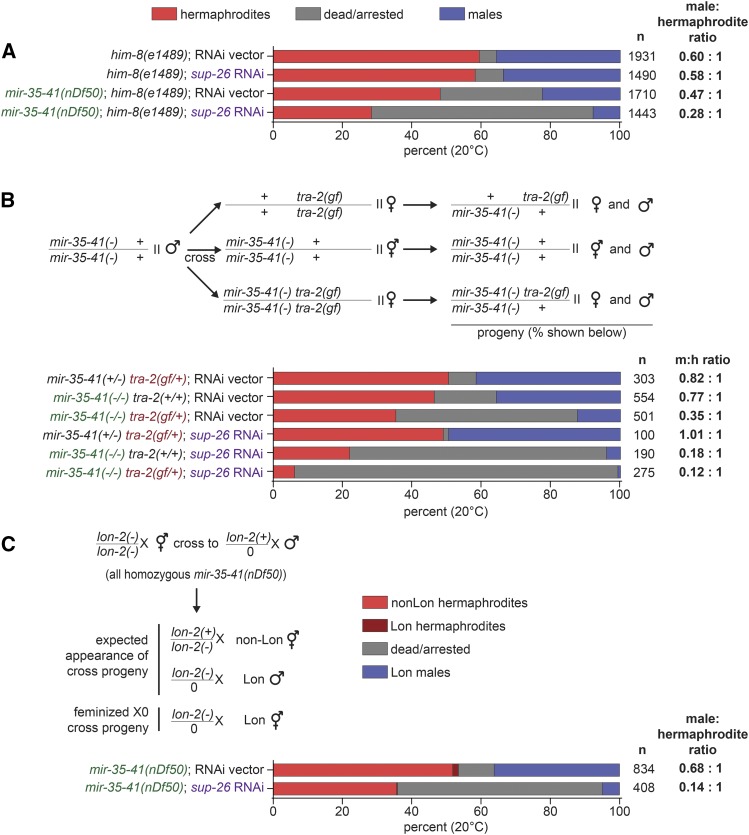
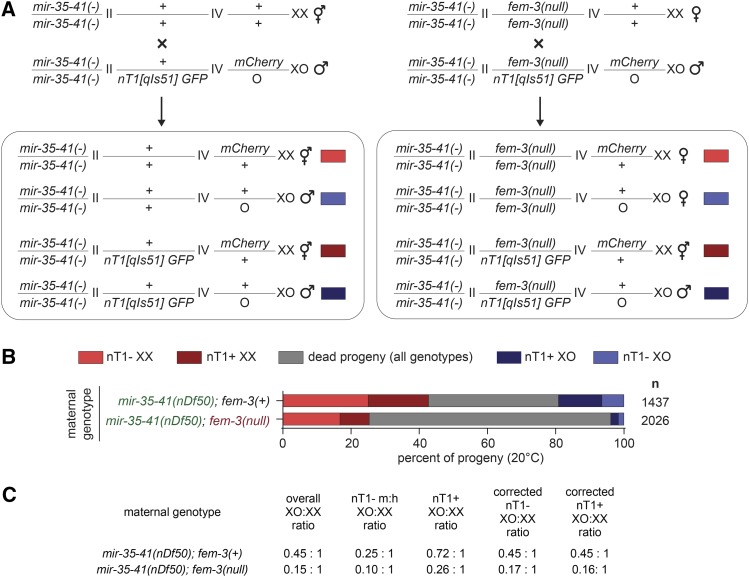
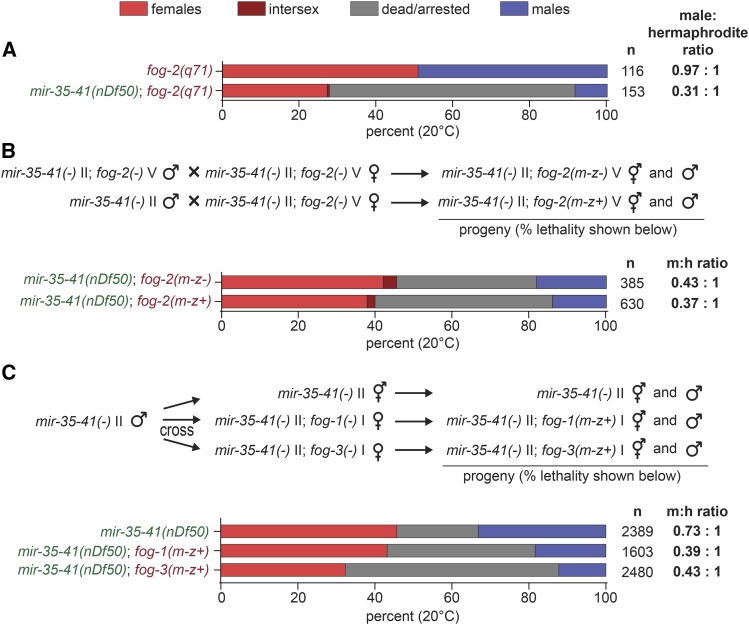
The germline sex determination pathway in C. elegans determines whether germ cells develop as oocytes or sperm, with no previously known effect on viability. The mir-35 family of microRNAs are expressed in the C. elegans germline and embryo and are essential for both viability and normal hermaphroditic sex determination, preventing aberrant male gene expression in XX hermaphrodite embryos. Here we show that combining feminizing mutations with partial loss of function of the mir-35 family results in enhanced penetrance embryonic lethality that preferentially kills XO animals. This lethal phenotype is due to altered signaling through the germline sex determination pathway, and maternal germline feminization is sufficient to induce enhanced lethality. These findings reveal a surprising pleiotropy of sperm-fate promoting pathways on organismal viability. Overall, our results demonstrate an unexpectedly strong link between sex determination and embryonic viability, and suggest that in wild type animals, mir-35 family members buffer against misregulation of pathways outside the sex determination program, allowing for clean sex reversal rather than deleterious effects of perturbing sex determination genes.
Keywords: Genetics of Sex; XO lethality; embryogenesis; germline sex determination; microRNAs; sex determination.
Copyright © 2019 Benner et al.
Figures
Similar articles
-
Sperm fate is promoted by the mir-44 microRNA family in the Caenorhabditis elegans hermaphrodite germline.Genetics. 2021 Mar 3;217(1):1-14. doi: 10.1093/genetics/iyaa006. Genetics. 2021. PMID: 33683352 Free PMC article.
-
A microRNA family exerts maternal control on sex determination in C. elegans.Genes Dev. 2017 Feb 15;31(4):422-437. doi: 10.1101/gad.290155.116. Epub 2017 Mar 9. Genes Dev. 2017. PMID: 28279983 Free PMC article.
-
A sensitized genetic background reveals evolution near the terminus of the Caenorhabditis germline sex determination pathway.Evol Dev. 2009 Jul-Aug;11(4):333-42. doi: 10.1111/j.1525-142X.2009.00340.x. Evol Dev. 2009. PMID: 19601967 Free PMC article.
-
Sex determination in the Caenorhabditis elegans germline.Adv Exp Med Biol. 2013;757:41-69. doi: 10.1007/978-1-4614-4015-4_3. Adv Exp Med Biol. 2013. PMID: 22872474 Review.
-
Maternal effects of microRNAs in early embryogenesis.RNA Biol. 2018 Feb 1;15(2):165-169. doi: 10.1080/15476286.2017.1402999. Epub 2017 Dec 8. RNA Biol. 2018. PMID: 29120257 Free PMC article. Review.
Cited by
-
Recent advances in understanding microRNA function and regulation in C. elegans.Semin Cell Dev Biol. 2024 Feb 15;154(Pt A):4-13. doi: 10.1016/j.semcdb.2023.03.011. Epub 2023 Apr 11. Semin Cell Dev Biol. 2024. PMID: 37055330 Review.
-
The multifaceted roles of microRNAs in differentiation.Curr Opin Cell Biol. 2020 Dec;67:118-140. doi: 10.1016/j.ceb.2020.08.015. Epub 2020 Nov 3. Curr Opin Cell Biol. 2020. PMID: 33152557 Free PMC article. Review.
-
In vivo CRISPR screening for phenotypic targets of the mir-35-42 family in C. elegans.Genes Dev. 2020 Sep 1;34(17-18):1227-1238. doi: 10.1101/gad.339333.120. Epub 2020 Aug 20. Genes Dev. 2020. PMID: 32820039 Free PMC article.
References
Publication types
MeSH terms
Substances
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources