

CCC- and WASH-mediated endosomal sorting of LDLR is required for normal clearance of circulating LDL
- PMID: 26965651
- PMCID: PMC4792963
- DOI: 10.1038/ncomms10961
CCC- and WASH-mediated endosomal sorting of LDLR is required for normal clearance of circulating LDL
Abstract
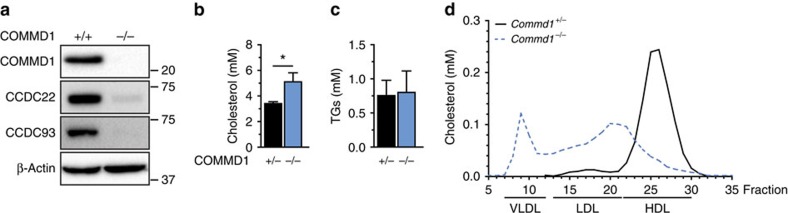
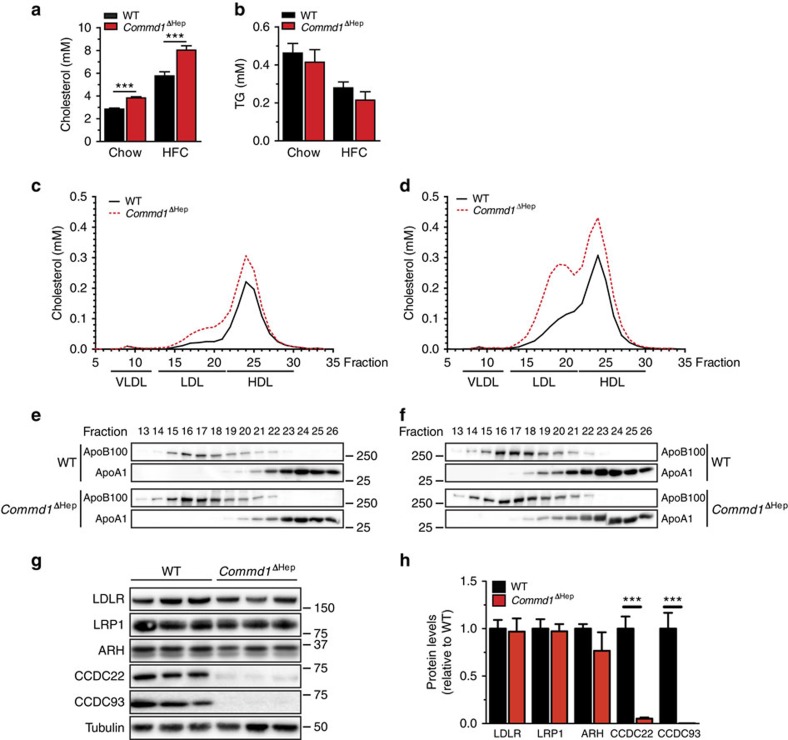
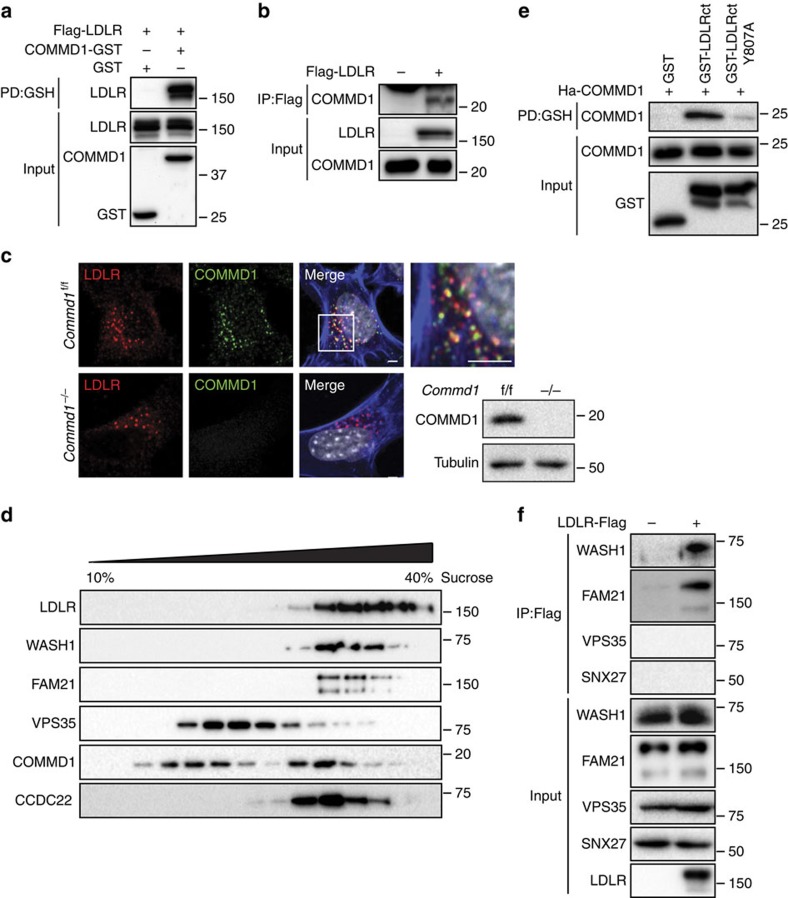
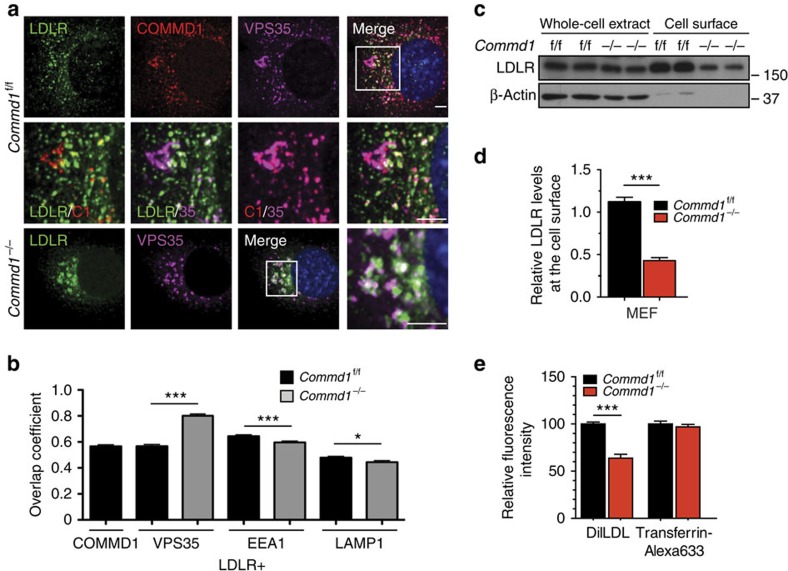
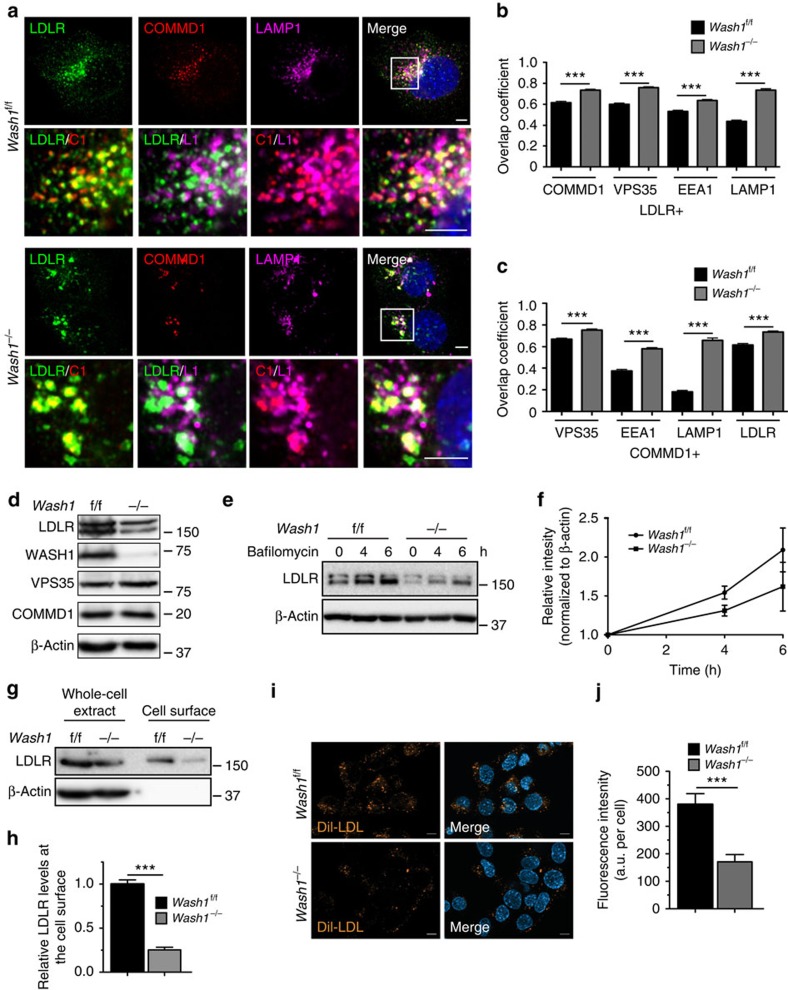
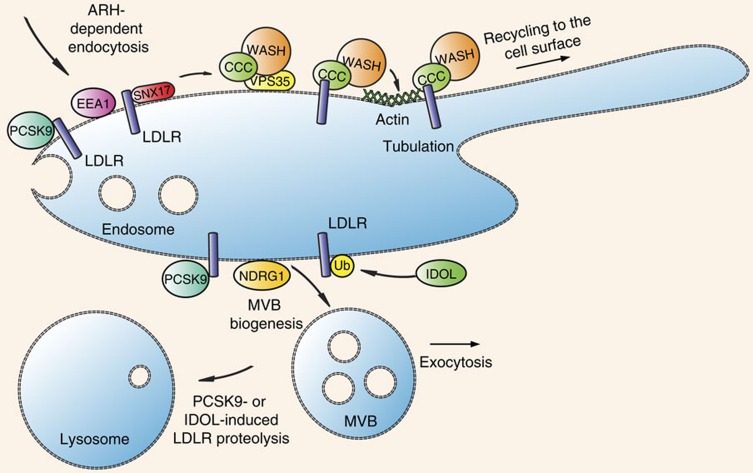
The low-density lipoprotein receptor (LDLR) plays a pivotal role in clearing atherogenic circulating low-density lipoprotein (LDL) cholesterol. Here we show that the COMMD/CCDC22/CCDC93 (CCC) and the Wiskott-Aldrich syndrome protein and SCAR homologue (WASH) complexes are both crucial for endosomal sorting of LDLR and for its function. We find that patients with X-linked intellectual disability caused by mutations in CCDC22 are hypercholesterolaemic, and that COMMD1-deficient dogs and liver-specific Commd1 knockout mice have elevated plasma LDL cholesterol levels. Furthermore, Commd1 depletion results in mislocalization of LDLR, accompanied by decreased LDL uptake. Increased total plasma cholesterol levels are also seen in hepatic COMMD9-deficient mice. Inactivation of the CCC-associated WASH complex causes LDLR mislocalization, increased lysosomal degradation of LDLR and impaired LDL uptake. Furthermore, a mutation in the WASH component KIAA0196 (strumpellin) is associated with hypercholesterolaemia in humans. Altogether, this study provides valuable insights into the mechanisms regulating cholesterol homeostasis and LDLR trafficking.
Figures
Similar articles
-
The COMMD Family Regulates Plasma LDL Levels and Attenuates Atherosclerosis Through Stabilizing the CCC Complex in Endosomal LDLR Trafficking.Circ Res. 2018 Jun 8;122(12):1648-1660. doi: 10.1161/CIRCRESAHA.117.312004. Epub 2018 Mar 15. Circ Res. 2018. PMID: 29545368
-
The hepatic WASH complex is required for efficient plasma LDL and HDL cholesterol clearance.JCI Insight. 2019 Jun 6;4(11):e126462. doi: 10.1172/jci.insight.126462. eCollection 2019 Jun 6. JCI Insight. 2019. PMID: 31167970 Free PMC article.
-
Normal sorting but defective endocytosis of the low density lipoprotein receptor in mice with autosomal recessive hypercholesterolemia.J Biol Chem. 2003 Aug 1;278(31):29024-30. doi: 10.1074/jbc.M304855200. Epub 2003 May 13. J Biol Chem. 2003. PMID: 12746448
-
News on the molecular regulation and function of hepatic low-density lipoprotein receptor and LDLR-related protein 1.Curr Opin Lipidol. 2017 Jun;28(3):241-247. doi: 10.1097/MOL.0000000000000411. Curr Opin Lipidol. 2017. PMID: 28301372 Free PMC article. Review.
-
The life cycle of the low-density lipoprotein receptor: insights from cellular and in-vivo studies.Curr Opin Lipidol. 2015 Apr;26(2):82-7. doi: 10.1097/MOL.0000000000000157. Curr Opin Lipidol. 2015. PMID: 25692346 Review.
Cited by
-
Eps15 Homology Domain Protein 4 (EHD4) is required for Eps15 Homology Domain Protein 1 (EHD1)-mediated endosomal recruitment and fission.PLoS One. 2020 Sep 23;15(9):e0239657. doi: 10.1371/journal.pone.0239657. eCollection 2020. PLoS One. 2020. PMID: 32966336 Free PMC article.
-
Use of plasma metabolomics to analyze phenotype-genotype relationships in young hypercholesterolemic females.J Lipid Res. 2018 Nov;59(11):2174-2180. doi: 10.1194/jlr.M088930. Epub 2018 Sep 28. J Lipid Res. 2018. PMID: 30266833 Free PMC article.
-
WASP family proteins: Molecular mechanisms and implications in human disease.Eur J Cell Biol. 2022 Jun-Aug;101(3):151244. doi: 10.1016/j.ejcb.2022.151244. Epub 2022 Jun 1. Eur J Cell Biol. 2022. PMID: 35667337 Free PMC article.
-
Phosphatidylinositol-(4,5)-Bisphosphate Regulates Plasma Cholesterol Through LDL (Low-Density Lipoprotein) Receptor Lysosomal Degradation.Arterioscler Thromb Vasc Biol. 2020 May;40(5):1311-1324. doi: 10.1161/ATVBAHA.120.314033. Epub 2020 Mar 19. Arterioscler Thromb Vasc Biol. 2020. PMID: 32188273 Free PMC article.
-
Proteomic and Biochemical Comparison of the Cellular Interaction Partners of Human VPS33A and VPS33B.J Mol Biol. 2018 Jul 6;430(14):2153-2163. doi: 10.1016/j.jmb.2018.05.019. Epub 2018 May 17. J Mol Biol. 2018. PMID: 29778605 Free PMC article.
References
-
- Harbour M. E., Breusegem S. Y. & Seaman M. N. J. Recruitment of the endosomal WASH complex is mediated by the extended ‘tail' of Fam21 binding to the retromer protein Vps35. Biochem. J. 442, 209–220 (2012) . - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
