

Spatial reconstruction of single-cell gene expression data
- PMID: 25867923
- PMCID: PMC4430369
- DOI: 10.1038/nbt.3192
Spatial reconstruction of single-cell gene expression data
Abstract
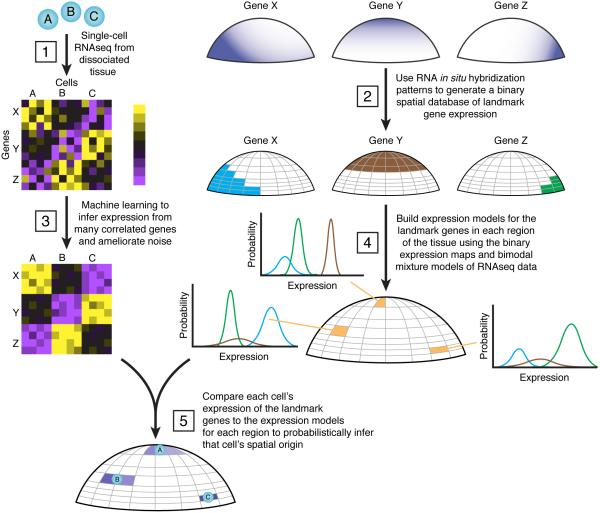
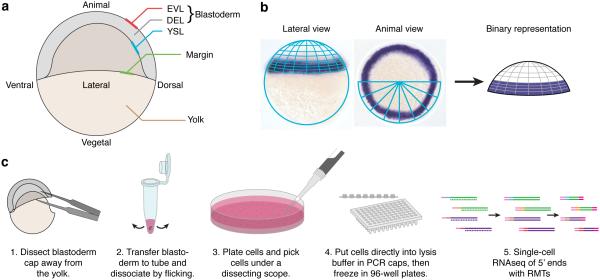
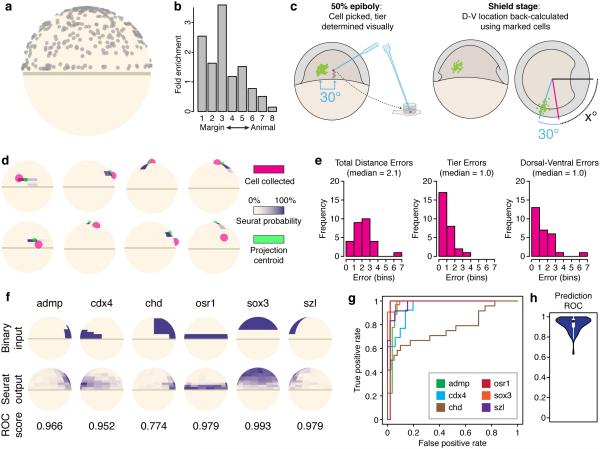
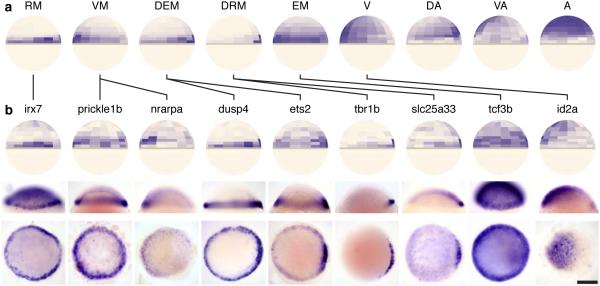
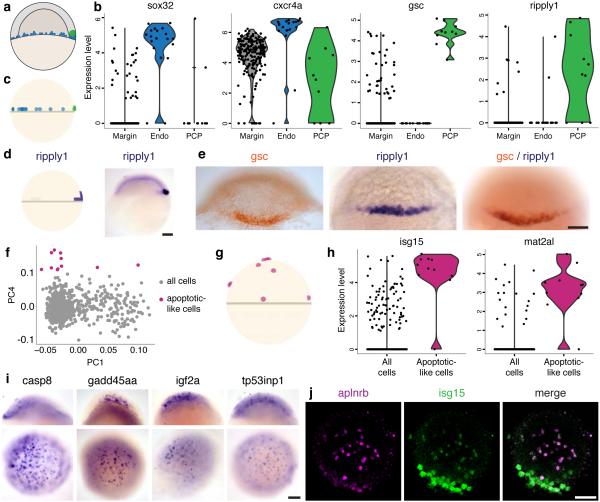
Spatial localization is a key determinant of cellular fate and behavior, but methods for spatially resolved, transcriptome-wide gene expression profiling across complex tissues are lacking. RNA staining methods assay only a small number of transcripts, whereas single-cell RNA-seq, which measures global gene expression, separates cells from their native spatial context. Here we present Seurat, a computational strategy to infer cellular localization by integrating single-cell RNA-seq data with in situ RNA patterns. We applied Seurat to spatially map 851 single cells from dissociated zebrafish (Danio rerio) embryos and generated a transcriptome-wide map of spatial patterning. We confirmed Seurat's accuracy using several experimental approaches, then used the strategy to identify a set of archetypal expression patterns and spatial markers. Seurat correctly localizes rare subpopulations, accurately mapping both spatially restricted and scattered groups. Seurat will be applicable to mapping cellular localization within complex patterned tissues in diverse systems.
Figures
Comment in
-
RNA: Putting transcriptomics in its place.Nat Rev Genet. 2015 Jun;16(6):319. doi: 10.1038/nrg3951. Epub 2015 May 7. Nat Rev Genet. 2015. PMID: 25948245 No abstract available.
Similar articles
-
Putting cells in their place.Nat Biotechnol. 2015 May;33(5):490-1. doi: 10.1038/nbt.3219. Nat Biotechnol. 2015. PMID: 25965757 No abstract available.
-
RNA Tomography for Spatially Resolved Transcriptomics (Tomo-Seq).Methods Mol Biol. 2019;1920:129-141. doi: 10.1007/978-1-4939-9009-2_9. Methods Mol Biol. 2019. PMID: 30737690
-
Spatial transcriptome profiling by MERFISH reveals subcellular RNA compartmentalization and cell cycle-dependent gene expression.Proc Natl Acad Sci U S A. 2019 Sep 24;116(39):19490-19499. doi: 10.1073/pnas.1912459116. Epub 2019 Sep 9. Proc Natl Acad Sci U S A. 2019. PMID: 31501331 Free PMC article.
-
Mapping Cellular Coordinates through Advances in Spatial Transcriptomics Technology.Mol Cells. 2020 Jul 31;43(7):591-599. doi: 10.14348/molcells.2020.0020. Mol Cells. 2020. PMID: 32507771 Free PMC article. Review.
-
Placing RNA in context and space - methods for spatially resolved transcriptomics.FEBS J. 2019 Apr;286(8):1468-1481. doi: 10.1111/febs.14435. Epub 2018 Mar 31. FEBS J. 2019. PMID: 29542254 Review.
Cited by
-
Aging disrupts blood-brain and blood-spinal cord barrier homeostasis, but does not increase paracellular permeability.Geroscience. 2024 Oct 30. doi: 10.1007/s11357-024-01404-9. Online ahead of print. Geroscience. 2024. PMID: 39476323
-
Inflammation impacts androgen receptor signaling in basal prostate stem cells through interleukin 1 receptor antagonist.Commun Biol. 2024 Oct 25;7(1):1390. doi: 10.1038/s42003-024-07071-y. Commun Biol. 2024. PMID: 39455902 Free PMC article.
-
A latent subset of human hematopoietic stem cells resists regenerative stress to preserve stemness.Nat Immunol. 2021 Jun;22(6):723-734. doi: 10.1038/s41590-021-00925-1. Epub 2021 May 6. Nat Immunol. 2021. PMID: 33958784
-
Single-cell immune repertoire sequencing of B and T cells in murine models of infection and autoimmunity.Genes Immun. 2022 Sep;23(6):183-195. doi: 10.1038/s41435-022-00180-w. Epub 2022 Aug 26. Genes Immun. 2022. PMID: 36028771 Free PMC article.
-
Loss of Gap Junction Delta-2 (GJD2) gene orthologs leads to refractive error in zebrafish.Commun Biol. 2021 Jun 3;4(1):676. doi: 10.1038/s42003-021-02185-z. Commun Biol. 2021. PMID: 34083742 Free PMC article.
References
-
- Schier AF. Genomics: Zebrafish earns its stripes. Nature. 2013;496:443–444. - PubMed
Publication types
MeSH terms
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
