

ABSTRACT
The gastrointestinal (GI) tract is considered the largest immunological organ in the body having a central role in regulating immune homeostasis. Contrary to earlier belief, the intestinal epithelial barrier is not a static physical barrier but rather strongly interacts with the gut microbiome and cells of the immune system. This intense communication between epithelial cells, immune cells and microbiome will shape specific immune responses to antigens, balancing tolerance and effector immune functions. Recent studies indicate that composition of the gut microbiome affects immune system development and modulates immune mediators, which in turn affect the intestinal barrier. Moreover, dysbiosis may favor intestinal barrier disruption and could be related to increased susceptibility to certain diseases. This review will be focused on the development of the intestinal barrier and its function in host immune defense and how gut microbiome composition throughout life can affect this role.
KEYWORDS: ageing, development, gut immunity, microbiota, mucosal barriers
The gastrointestinal tract and its role in host defense
The gastrointestinal (GI) tract is the largest interface in the body that is in contact with the external environment, fulfilling the function of food processing and allowing the survival of commensal symbiotic microbes while eliminating pathogens. The GI tract is therefore, a major line of defense in which epithelial cells provide a physical barrier and work in concert with immune and stromal cells to fight off pathogens and limit their direct contact with the epithelium.1 The immune system of the GI tract is constantly challenged with antigens from the lumen and therefore must be able to distinguish which antigens should be tolerated or not (self-antigens, food, symbiotic microbes vs. pathogens, toxins). The epithelium is constituted of a single layer of different intestinal epithelial cells (IECs) that strongly participates in innate immunity (Fig. 1). Endocrine cells and stem cells at the base of the intestinal crypts give rise to IECs with specialized function (enterocytes, goblet cells, Paneth cells and enteroendocrine cells). Enterocytes and Paneth cells (PC) produce antimicrobial peptides, such as alpha-defensins, lysozyme C, phospholipases, C-type lectin, and regenerating islet-derived 3-gamma (RegIIIγ),2 that are important to keep pathogens under control. Goblet cells secret mucins that lubricate and protect the epithelial intestinal surface, as well as, participate as antigen presenting cells (APCs) delivering luminal antigens to CD103+ dendritic cells (DCs), which promote the development of regulatory T cells (Tregs).3 Furthermore, the continuous regulated turnover of IECs serves as a defense mechanism by helping to prevent pathogen attachment to the intestinal wall and colonization of the gut.4,5 Another essential piece of the intestinal barrier machinery are tight junction complexes, consisting of claudins, occludins, zonula occludens and junction adhesion molecules, which create a seal between neighboring IECs and impede entry of pathogens while regulating permeability to water, ions and nutrients.5,6 Under normal circumstances there is a dynamic regulation of tight junction components, however sustained inflammation or infections can lead to dysregulation in the expression of adhesion molecules, leading to barrier breach and entry of microbes.1,6
Figure 1.

Schematic representation of mucosa, villi, crypts of Lieberkühn and cells of the small intestine. The intestinal lining of the lower intestine is highly folded to maximize absorption and contains finger-like mucosal projections that form structures called villi, in between the villi downward invaginations, called crypts of Lieberkuhn, extend down to the muscularis mucosae. Underlying the epithelium, the lamina propria, harbors dendritic cells, important antigen-presenting cells, which regulate humoral and cellular gut immunity. The muscularis externa layer contains two layers of smooth muscle that enable continuous peristaltic activity of the small intestine. On the left, the epithelium of a crypt and part of a villus are represented and different epithelial cells can be identified: enterocytes, tall columnar absorptive cells with ‘brush-like border’ on the apical surface, called microvilli; goblet cells, which secrete mucin, for lubrication of the intestinal contents and protection of the epithelium; enteroendocrine cells that secrete various gut hormones; stem cells that lie near the base of the crypt and give rise to the specialized epithelial cells; above the stem cells are transit amplifying cells; and Paneth cells, which have a defensive function secreting antimicrobial molecules into the lumen.
In general, it is estimated that the GI tract can harbour up to 70% of the body´s lymphocytes population, making it the largest immunological organ in the body7 (Fig. 1). Under the epithelium, the lamina propria (LP) harbours dendritic cells (DCs), which are important APCs,8 and the gut-associated lymphoid tissue (GALT), that includes Peyer´s patches (PP), LP-lymphocytes and intraepithelial lymphocytes (IELs). The PP, more frequent in the ileum where bacterial load is greater, are important induction sites that contain all the immune-competent cells necessary to induce antigen-specific responses.9 They also have a specialized follicle epithelium of microfold (M) cells, capable of sampling and transporting antigens from lumen to the underlying immune machinery.10 M cells can be considered double-edged swords because they mediate the passage of the antigens to the mucosal lymphoid tissue for initiation of immune responses but can also serve as an entry point for microbes.10 Finally, the adaptive immune system contributes to intestinal barrier defense by secreting effector factors, such as immunoglobulins (Ig), into the intestinal lumen to combat pathogen attachment and invasion of mucosal tissue.11
In recent years, microbiota has been linked to host immunity. Studies have shown that not only gut microbiota is important for the metabolism of essential nutrients for the organism but it also plays a crucial role in gut development and is necessary for the development of a fully functional immune system.12,13 In the next sections, the influence of microbiota in the development of the intestinal barrier and the immune system will be discussed.
Microbiota shapes the development of the immune system and host immune responses
The skin and mucosa are covered by a wide range of microorganisms, including fungi, virus, parasites and bacteria, a large percentage colonizes the GI tract and therefore are called gut microbiota.12,14,15 The human GI tract contains approximately 100 trillion bacteria and renders this location the primary site of interactions between microbes and the host immune system.12,16 Millions of years of co-evolution with our microbiota have ensured a beneficial mutualistic relationship, in which the microbiota contributes to many physiological functions of the host, while in return the host offers nourishment and habitat. Beyond aiding in the digestion and fermentation of food that is important for production of certain vitamins, the microbiota is also essential in the defense against pathogens once they compete for nutrients and adhesion sites, some even actively eliminating competition by secreting antimicrobial peptides.14,15 A stable microbiota together with mucus layers are essential to prevent pathogenic bacteria from causing host infections.
Recently, experiments conducted in germ-free (GF) animals demonstrated that microbiota colonization in early life is necessary for optimal development of the immune system. In the absence of microbiota, intestinal mucosal immunity is underdeveloped and animals present smaller mesenteric lymph nodes, PP and reduced numbers of immune cells such as IgA-producing plasma cells, CD4+ LP T-cells and intraepithelial αβ T-cell receptor CD8+ cells, resulting in a weakened capacity to fight off pathogenic bacteria.17 Earlier studies had already suggested that housing conditions affecting microbiota colonization led to immune alterations, for example, absence of microbiota generated GF mice with unstructured spleen and lymph nodes with disorganized B- and T-zones and reduced serum levels of IgG in comparison to mice raised in conventional housing.18,19 Furthermore, lack of intestinal bacteria in GF animals, decreased functional CD4+CD25+ Tregs.16 Littman et al.20 demonstrated that the balance between pro-inflammatory interleukin (IL)-17-producing effector T helper (Th17) cells and Foxp3+ Tregs in the gut, required signals from intestinal bacteria and was dependent on the composition of the gut microbiota.
One of the possibilities of host physiological regulation by microbiota is their production of an extremely diverse metabolite repertoire that results from anaerobic fermentation of leftover food in the digestive tract. The major products that results from bacterial fermentation of fibers in the colon are short-chain fatty acids (SCFAs), including acetic acid, butyric acid and propionic acid, which gain access through the intestinal epithelia and are able to interact with host cells, thus influencing immune responses and disease risk.21 Not only are SCFAs important energy sources for the gut microbiota itself but also for IECs and have diverse regulatory functions in host physiology and immunity, in general being regarded as beneficial metabolites with anti-inflammatory properties.22 Microbial gut composition and fiber availability in diet influence SCFA concentration in the colon. A diet rich in fibers can favor the presence of bacteria capable of cellulose and xylan hydrolysis, including members of the genus Prevotella, Xylanibacter and the butyrate-producing Faecalibacterium prausnitzii (F. prausnitzii).22,23 It has also been hypothesized that high abundance of F. prausnitzii along with other SCFA-producing bacteria could protect the host from inflammation and noninfectious colonic diseases. To that effect, reports have correlated a low abundance of F. prausnitzii with Crohn´s Disease (CD) and others inflammatory bowel diseases (IBDs).24,25 SCFAs are inhibitors of histone deacetylases (HDACs) that tend to promote a tolerogenic, anti-inflammatory cell phenotype that is crucial for maintaining immune homeostasis.13 A strong example of microbiota influence on the immune system via epigenetic control is the regulation of Treg differentiation by butyrate (a SCFA). Naïve CD4+ T cells cultured in Treg differentiation conditions together with butyrate, presented enhanced acetylation of histone H3 in lysine 27 (H3K27) at the Foxp3 promoter and CNS1 and CNS3 enhancer, thus, leading to epigenetic modifications that increased Foxp3 induction and an enhanced regulatory capacity of Tregs.26 Furthermore, SCFAs also enhance defense mechanisms by fortifying IECs barrier function. In vivo, it was shown that GF mice colonized with SCFA-producing Bacteroides thetaiotaomicron or F. prausnitzii induced goblet cell differentiation and mucus production.27 In vitro, intestinal epithelial goblet cells increased their transcription of mucin genes in response to SCFAs.28,29 Similarly, different SCFAs facilitate tight junction assembly in IECs and colonization with a strain of Bifidobacterium longum that produces high levels of acetate conferred protection against lethal enteropathogenic Escherichia coli O157:H7 infection. This suggests that SCFAs can enhance IEC integrity and inhibit the translocation of lethal toxins from the gut lumen into the systemic circulation.30-32
It is not clear exactly how microbial composition regulates immune homeostasis but some studies show that the presence of specific bacteria species can shift immune responses by favoring the development of certain subtypes of lymphocytes, for example, segmented filamentous bacteria (SFB) induce IL-17 and IL-22 production and favor the generation of Th17 cells in mice,33 while microbiota reconstitution of GF mice with Clostridium spp, altered Schaedler flora and the bacteria Bacteroides fragilis promote the accumulation of IL-10+ Tregs in the colon of this animals.34-36 Furthermore, it appears that recognition of microbial stimuli is important for immune regulatory mechanisms, as it was shown that deletion of Myeloid differentiation primary response gene 88 (Myd88), a protein that acts as an adaptor of multiple innate immune receptors, in the nonobese (NOD) diabetic mouse, a model of spontaneous type 1 diabetes, resulted in less severe diabetes and an altered microbial composition, while depletion of microbiota led to development of robust diabetes.37
Currently, there are schools of thought supporting the idea that the microbiome and the intestinal immune system are key in sustaining not only local but systemic immune regulation and that dysbiosis could favor effector immune responses that trigger autoimmune disorders.38 Considering that a stable microbiota is important to maintain a well-balanced immune system, it is worrisome that antibiotics are one of the most commonly prescribed drugs for children. Antibiotics disrupt the delicate ecosystem in the gut of the young infant and could possibly augment the risk of autoinflammatory diseases later in life.39-41 The indiscriminate depletion of commensal bacteria following antibiotic intake results in vacating niches, which can increase host vulnerability to excessive colonization by opportunistic pathogens and create dysbiosis. Supporting this hypothesis, studies determined that the use of antibiotics in young mice resulted in altered microbiota, induced a shift towards pro-inflammatory immune responses and increased risk of inflammatory disease.42 In new-born mice, treatment with antibiotics depleted bacteria of the Clostridia class, diminished production of IL-22 by RORγt+ innate lymphoid cells (ILCs) and T-cells that reduced the access of the antigen to the bloodstream and prevented the allergens to cross the intestinal epithelial layer, resulting in an enhanced sensitization to food allergens.43 Low-dose penicillin in early life caused transient perturbations in the microbiota with persistent sustained metabolic alterations.44 In addition, the presence of SFB has been linked to some autoinflammatory diseases, such as exacerbating autoimmune encephalitis and murine arthritis models.45,46 Some case reports provide evidence that SFB could be detected at certain inflammatory sites both in ulcerative colitis (UC) and Chron´s disease (CD) patients, while depletion of SFB by penicillin decreased Th17 and susceptibility to Dextran Sodium Sulfate (DSS)-induced colitis in mice.47 On the other hand, SFB colonization could be important for the defense against bacterial pathogens, for instance, antibiotic use reduced SFB and Th17 cell numbers, decreasing resistance against intestinal pathogen Citrobacter rodentium (C. rodentium) infection. In contrast, colonization of mice with SFB conferred higher resistance to C. rodentium and was correlated with increased expression of genes associated with inflammation and antimicrobial defenses.33
Given the ever-growing data appointing our microbiome as a master regulator of various systems which must collaborate to ensure human health, scientists have joined forces and mounted consortiums such as The Human Microbiome project launched in 2008 with the aim to characterize and identify the composition of human microbiota and its role in health and disease.48 Despite these efforts, it has been very difficult to determine what defines a healthy microbiota. This is in part because individual (epi)genetic variability is very high, thus pinpointing how the loss or gain of specific bacteria will affect individuals is very complex. Also, the gut microbiome composition changes throughout life and in order to design effective therapies to prevent disease it is important to determine specific time-frames to intervene in microbial gut composition. Many studies showed that developmental abnormalities caused by absent microbiota can be reversed by restoring intestinal microbial colonization only at a very young age. For example, the microbiota seems to have a role in maintaining the regulation of the T-helper 2 (Th2) population which is important for oral tolerance induction, reconstitution of GF mice with Bifidobacterium infantis could re-establish Th2-mediated responses and oral tolerance induction only when performed in neonates and not in older animals.49 Thus, understanding how specific microbial colonization regulates intestinal barrier immune responses would possibly enable medical interventions in microbiota to aid in the prevention or treatment of certain illnesses.
Early-life microbiota affects immunity later in life
During infancy, the diversity and variation in gut microbiota is high and tends to become less diverse and stabilize later in life50 (Fig. 2). Several environmental factors modulate the development of a child´s microbiota, including diet, use of antibiotics during perinatal period, mode of delivery at birth, breast-feeding and infections51 (Fig. 2). Considering that microbiota influences the development of the immune system, it is important to comprehend how early-life microbiota composition can be a risk factor for numerous diseases, including allergy and autoimmunity.52,53
Figure 2.
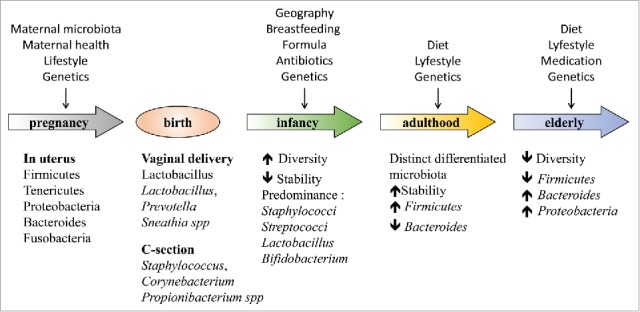
Changes in human microbiota throughout life. The uterus is not a sterile environment, studies have found bacteria in placentae,95,96 fetal membranes,97 umbilical cord blood98 and meconium.99,100 Colonization of the infant gut will depend on the mode of birth, vaginally-delivered and Cesarean section (C-section) children acquire distinct bacterial communities.105,106 In young children, the composition of the gut microbiota often varies, is very diverse and less stable, with age gut microbiota becomes more stable. In old age, the gut microbiota alters, it is less diverse compared to younger age and presents reduction in short chain fatty acid-producing bacteria and increase in gram-negative LPS-secreting bacteria.58,144
Pregnancy also affects the immunological system of mothers and infants. Pregnant mothers develop a Th2 immunological bias that is more intense at the maternal-fetal interface and is controlled by the mother's hormones.54 Serum from pregnant women have higher levels of IL-4 and IL-10 regulatory cytokines, and reduced levels of pro-inflammatory interferon (INF)-γ and IL-2.55 Moreover, trophoblasts and endometrial gland cells produce the chemokine C-C Motif Chemokine Ligand (CCL)17 that promotes the infiltration of CCR4+ Th2-type cells into the human decidua during early pregnancy.56 Local production of Th2 cytokines, such as IL-4, IL-10 and IL-13, inhibits Th1 cell development and Th1 cytokine production thereby protecting the fetus and preventing rejection during early gestation.57 There is growing evidence that the development of the fetal immune system is actively and passively controlled by maternal immunity, and that the consequences of this interaction have a strong impact on immunity and immune disorders later in life, such as development of allergies.58-60
In recent years, the idea of a sterile uterus has been overturned by culture-based and independent studies identifying the presence of bacteria in placentae,61,62 fetal membranes,63 umbilical cord blood64 and meconium.65,66 DNA of Bifidobacterium and Lactobacillus rhamnosus, two commensals found in the adult gut, was found in placentae samples.61,67 It is proposed that horizontal transfer of bacterial DNA from mother to fetus occurs via the placenta´s bloodstream, as Jiménez et al.64,65 showed by detecting Enterococcus faecium in umbilical cord blood and the meconium in mice previously orally inoculated with the bacteria. Another study reported the presence of low quantities of bacteria in human placenta by identifying a nonpathogenic microbiota niche (Firmicutes, Tenericutes, Proteobacteria, Bacteroides and Fusobacteria), which is similar to microbiota present in the oral cavity.61,62 In addition, experimental murine models demonstrated that maternal treatment with injected and/or aerosolized lipopolysaccharides (LPS) or intranasal bacteria (Acinetobacter lwoffi) or given as probiotic supplementation (L. rhamnosus) during gestation attenuated allergic sensitization and airway inflammation in the offspring, induced mucosal tolerance and suppressed the production of allergen-induced IgE, eosinophilic airway inflammation and airway reactivity.68-70
In mammals, infants receive a different bacterial inoculum depending on their mode of delivery which determines differential infant gut microbial colonization. Vaginally delivered children acquire bacterial communities present in their own mother's vaginal microbiota (Lactobacillus, Prevotella, or Sneathia spp.). In contrast, Cesarean section (C-section) infants harbour bacterial communities like those found on the mother´s skin and environmental surfaces (Staphylococcus, Corynebacterium, and Propionibacterium spp.).71,72 In an experimental setting, it was demonstrated that the mode of delivery impacted tolerance in the delivered pups. Epithelial tolerance was only observed in vaginally-delivered and not C-section pups. IECs from vaginally-delivered neonates were resistant to LPS and presented spontaneous activation of nuclear factor (NF)-κB together with a post-transcriptional down regulation of interleukin 1 receptor-associated kinase 1 (IRAK-1), essential for epithelial Toll like receptor (TLR)4 signaling in vivo.73 Tolerance is critical for the development of a stable homeostasis between host and microbiome. Thus, this neonate colonization seems to regulate the immune system early on.
While the infant is growing, diet has a great influence on the composition of the microbiota. Children that receive formula or breast milk will receive distinct nutrients, microbial and immune components. Breast milk is important for the infant´s immune defenses as it contains IgA, lactoferrin and defensins, protecting against infections and contributing to the maturation of the immune system in the first days of life.74,75 Researchers have also found that human breast milk contains neurotrophic factors and cytokines that influence enteric neuron survival and neurite outgrowth.76 Importantly, contrary to common belief, human milk is not sterile, containing commensal, mutualistic and/or potentially probiotic bacteria to offspring. The composition of bacteria found in breast milk can vary with geography and maternal diet but several studies have revealed predominant presence of Staphylococci, Streptococci, Lactobacillus and Bifidobacterium.77 Data collected so far suggest that lack of breast-feeding could increase health risks in infants. For example, premature infants that did not receive breast milk presented an increased risk of necrotizing enterocolitis (NEC).78 Also, absence of breastfeeding in infants is associated with an increased incidence of many diseases such as infectious morbidity, gastroenteritis, pneumonia, childhood obesity, type 1 and type 2 diabetes, leukemia, and sudden infant death syndrome (SIDS).79
Microbiota and the intestinal barrier
Since birth, the microbiota begin the colonization of the GI tract and participate in many host physiological processes including intestinal barrier immunity.80 In mice, depletion of microbiota affected IECs, resulted in altered patterns of microvilli formation and decreased cell renewal.81 Also, the enteric nervous system (ENS) development is greatly influenced by microbiota. GF mice and antibiotic–treated animals present a reduced number of enteric neurons, alterations in neurotransmitter expression, delayed gastric emptying and intestinal transit.82,83 In addition, specific strains of bacteria were described to affect small intestine myoelectrical activity and intestinal transit.84 The communication of the ENS and the microbiota is not “one-way” because ENS inputs also influence microbiota composition. Wiles and collaborators showed that gut motility is important for the establishment of bacterial communities during development. Their study demonstrated that mutation of the ret locus, which is associated with the intestinal motility disorder Hirschsprug disease in humans, allowed co-habitation of usually competitive bacterial species in the gut, in contrast to wild types in which these competitive species differed in abundance.85 It seems that this dynamic environment is a prerequisite for a balanced microbial community and immune system. This is supported by the observation that ENS zebrafish mutants develop microbiota-dependent inflammation due to a surfeit of pro-inflammatory bacterial strains in detriment of anti-inflammatory strains. Transfer of microbiota from the ENS mutant transmitted this inflammation to individuals with a normally functioning ENS and interestingly, transferring an anti-inflammatory strain to mutants restored ENS functionality and reduced the inflammation.86
Mucus production is also pivotal for the organism´s defense as it forms a physical barrier separating bacteria from the host and avoids breach of the intestinal barrier. Failure of this protective barrier to separate bacteria from the epithelium can lead to disease and can be observed in murine colitis models and human IBDs.87 Compared to rodents, the human colon has an even thicker inner mucus layer separating bacteria from the tissue. In UC patients, this mucosal barrier is compromised and the bacteria can be seen in direct contact with the epithelium in patients with the active disease.87 Until recently it was not known if microbiota had any direct role in host mucus production. Now studies show that the presence of bacteria influences mucus quality. A Swedish study compared mucus of GF-mice versus conventionally-raised (conv)-mice. Their study showed that although the colon mucus organization of GF mice was similar to conv mice, the GF inner mucus layer was more penetrable to bacteria-sized beads.88 Other researchers analyzed the colon mucus barrier in different wild-type C57BL/6 mouse colonies housed and bred separately in the same specific pathogen-free (SPF) mouse facility. They observed that genetically identical animals housed in the same facility but kept in different rooms can have rather distinct microbiotas and mucus barrier structures. Also, they identified that mice with more impenetrable mucus layer had increased amounts of Erysipelotrichi, whereas mice with a penetrable mucus layer had higher concentrations of Proteobacteria and candidate division TM7 bacteria in the distal colon mucus.89 It is still unclear how microbial composition can regulate mucus properties but it is possible that microbe stimulation of intestinal cells can increase production of proteins involved in host responses and of core mucus proteins.
In summary, early microbiota colonization is an important event to establish the successful maturation and homeostasis of the intestinal barrier immune defense. Current data demonstrates a complex intercommunication between microbiota, immunity and the intestinal barrier, which must work together to confer protection against pathogens while maintaining tolerance and balance.
Ageing changes microbiota and intestinal barrier immunity
Improvement in sanitary conditions and health care have contributed to extend the life-expectancy and increase the ageing population worldwide. It is therefore important to know how ageing affects intestinal barrier immunity. In general, elderly individuals are considered at more risk of acquiring infections and other diseases, including many GI tract disorders. Nevertheless, little is known on how ageing affects epithelial cell immune function which is critical for host defense.
Amongst the most common GI tract disorders that afflict the ageing population is chronic constipation, which affects more than 50% of elderly people in home care so that up to 74% of this group has to use laxatives daily.90 Evidence suggests that motility is impaired in some GI tract regions during ageing but it remains to be determined to which extent failing smooth muscle cells or impaired ENS activity is to blame.91 Older mice present higher smooth muscle contractility when electrically stimulated but they also present increased thickness of muscle layers, which could impair bowel movement.92 Several different studies indicate there are changes in cellular proteins and mechanisms that regulate the contractile properties of the ageing intestinal smooth muscle such as, dysregulation of calcium signaling, changes in the signaling pathways that regulate the phosphorylation of myosin light chain, mitochondrial structure abnormalities and apoptosis of smooth muscle cells.91,93,94 On the hand, enteric neural loss during ageing seems to be variable and study results vary, some do not find significant loss while others report up to 50% neural loss.91,95 Other evidence of age-related neurodegeneration has also been described: swollen and dystrophic nerves in the gut, degenerating nerve fibers in rat small intestine and accumulation of lipofuscin in enteric neurons.96-98 There is growing evidence which suggest that despite the continuous self-renewal of intestinal epithelium, there is an accumulation of age-related molecular changes in long-lived stem/progenitor cells which could contribute to GI tract malfunction.99 Moreover, ageing myenteric neurons also display a senescence-associated phenotype.100
Age-related changes in the immune system also seem to affect cytokine production and ageing is related to the excessive production of ‘‘geriatric’’ pro-inflammatory cytokines, such as IL-6, tumor necrosis factor (TNF)-α and IL-1β.101 A possible reason for this alteration could be that ageing affects IEC-function once IECs are an important source of cytokines that regulate effector function of various cell types in the intestinal mucosa such as DCs and Treg cells.102,103 Furthermore, reactive oxygen species (ROS) have been detected in the ageing ENS, for example, higher production of ROS is found in myenteric neurons in the ileum of old rats compared to young animals.104 These increased levels of inflammatory mediators may have a direct impact on the aged gut, such as cytokine-induced dysregulation of tight junction barrier resulting in increased gut permeability (‘‘leaky gut’’).
Recently, it was demonstrated that changes in the microbiota occur during ageing and they correlate with health status and diet91 (Fig. 2). One of these age-related changes in composition of gut microbiota is an increase in the abundance of LPS-secreting Gram-negative bacteria, like Bacteroides, Proteobacteria and other pathogens.22,105 The rise in bacteria secreting LPS, which may act as endotoxin, could cause inflammation in gut. In addition, another factor that could contribute to increased gut-related diseases in the elderly is a decrease in intestinal SCFA concentrations, especially acetate, butyrate, and propionate and a reduction in Firmicutes phylum, whose primary metabolic end product is butyrate, compared to young subjects.22,106
Better understanding on how specific populations of bacteria regulate cytokine production is necessary to design future therapies in the elderly. In order for ageing population to maintain a healthy microbiome composition, it could be interesting to administer beneficial bacteria and target harmful bacteria.
Conclusion: Microbial diversity and implications for health
The gut and its epithelial barrier have become hot topics in the scientific community. Findings in the recent years, have demonstrated that the IECs are not merely static barriers that separate the external environment from our organisms. On the contrary, IECs engage in a complex-dynamic crosstalk with the intestinal immune system and microbiota. Moreover, early colonization of gut by microbes affects the development of the immune system and the microbial diversity throughout life varies and seems to affect susceptibility to some diseases. The dynamic interface of microbiota and the host´s environment, age, diet and health status complicate the identification of the role of specific bacteria in immune regulation. However, integrating all these observations is necessary to develop precise diagnostics and may indicate the necessity of more individualized designed therapies that are based on age and the microbiome.
Disclosure of potential conflicts of interest
No potential conflicts of interest were disclosed.
Acknowledgments
We thank Prof. Dr. Shaker Chuck Farah (Department of Biochemistry, Institute of Chemistry, University of Sao Paulo) for his constructive reading and comments. We also thank the CAPES-PNPD program for the postdoctoral fellowship of Dr. Tatiana Takiishi, and FAPESP for the doctoral fellowship of Camila Ideli Morales Fénero.
References
- [1].Odenwald MA, Turner JR. The intestinal epithelial barrier: a therapeutic target? Nat Rev Gastroenterol Hepatol. 2017;14:9-21. doi: 10.1038/nrgastro.2016.169. PMID:27848962 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [2].Pott J, Hornef M. Innate immune signalling at the intestinal epithelium in homeostasis and disease. EMBO Rep. 2012;13:684-98. doi: 10.1038/embor.2012.96. PMID:22801555 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [3].McDole JR, Wheeler LW, McDonald KG, Wang B, Konjufca V, Knoop KA, Newberry RD, Miller MJ. Goblet cells deliver luminal antigen to CD103+ dendritic cells in the small intestine. Nature. 2012;483:345-9. doi: 10.1038/nature10863. PMID:22422267 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [4].Cliffe LJ, Humphreys NE, Lane TE, Potten CS, Booth C, Grencis RK. Accelerated intestinal epithelial cell turnover: a new mechanism of parasite expulsion. Science. 2005;308:1463-5. doi: 10.1126/science.1108661. PMID:15933199 [DOI] [PubMed] [Google Scholar]
- [5].Ramanan D, Cadwell K. Intrinsic Defense Mechanisms of the Intestinal Epithelium. Cell host & microbe. 2016;19:434-41. doi: 10.1016/j.chom.2016.03.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [6].Marchiando AM, Graham WV, Turner JR. Epithelial barriers in homeostasis and disease. Annu Rev Pathol. 2010;5:119-44. doi: 10.1146/annurev.pathol.4.110807.092135. PMID:20078218 [DOI] [PubMed] [Google Scholar]
- [7].Pabst R, Russell MW, Brandtzaeg P. Tissue distribution of lymphocytes and plasma cells and the role of the gut. Trends in immunology. 2008;29:206-8; author reply 9–10. doi: 10.1016/j.it.2008.02.006. PMID:18394963 [DOI] [PubMed] [Google Scholar]
- [8].Uematsu S, Fujimoto K, Jang MH, Yang BG, Jung YJ, Nishiyama M, Sato S, Tsujimura T, Yamamoto M, Yokota Y, et al.. Regulation of humoral and cellular gut immunity by lamina propria dendritic cells expressing Toll-like receptor 5. Nature Immunol. 2008;9:769-76. doi: 10.1038/ni.1622. [DOI] [PubMed] [Google Scholar]
- [9].Mowat AM, Agace WW. Regional specialization within the intestinal immune system. Nature reviews Immunology. 2014;14:667-85. doi: 10.1038/nri3738. PMID:25234148 [DOI] [PubMed] [Google Scholar]
- [10].Mabbott NA, Donaldson DS, Ohno H, Williams IR, Mahajan A. Microfold (M) cells: important immunosurveillance posts in the intestinal epithelium. Mucosal Immunol. 2013;6:666-77. doi: 10.1038/mi.2013.30. PMID:23695511 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [11].Macpherson AJ, Geuking MB, Slack E, Hapfelmeier S, McCoy KD. The habitat, double life, citizenship, and forgetfulness of IgA. Immunological reviews. 2012;245:132-46. doi: 10.1111/j.1600-065X.2011.01072.x. PMID:22168417 [DOI] [PubMed] [Google Scholar]
- [12].Caricilli AM, Castoldi A, Camara NO. Intestinal barrier: A gentlemen's agreement between microbiota and immunity. World journal of gastrointestinal pathophysiology. 2014;5:18-32. doi: 10.4291/wjgp.v5.i1.18. PMID:24891972 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [13].Rooks MG, Garrett WS. Gut microbiota, metabolites and host immunity. Nature reviews Immunology. 2016;16:341-52. doi: 10.1038/nri.2016.42. PMID:27231050 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [14].Moens E, Veldhoen M. Epithelial barrier biology: good fences make good neighbours. Immunology. 2012;135:1-8. doi: 10.1111/j.1365-2567.2011.03506.x. PMID:22044254 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [15].Kamada N, Seo SU, Chen GY, Nunez G. Role of the gut microbiota in immunity and inflammatory disease. Nat Rev Immunol. 2013;13:321-35. doi: 10.1038/nri3430. PMID:23618829 [DOI] [PubMed] [Google Scholar]
- [16].Hara N, Alkanani AK, Ir D, Robertson CE, Wagner BD, Frank DN, Zipris D. The role of the intestinal microbiota in type 1 diabetes. Clin Immunol. 2013;146:112-9. doi: 10.1016/j.clim.2012.12.001. PMID:23314185 [DOI] [PubMed] [Google Scholar]
- [17].Sommer F, Backhed F. The gut microbiota–masters of host development and physiology. Nat Rev Microbiol. 2013;11:227-38. doi: 10.1038/nrmicro2974. PMID:23435359 [DOI] [PubMed] [Google Scholar]
- [18].Bauer H, Horowitz RE, Levenson SM, Popper H. The response of the lymphatic tissue to the microbial flora. Studies on germfree mice. AM J Pathology 1963;42:471-83. [PMC free article] [PubMed] [Google Scholar]
- [19].Benveniste J, Lespinats G, Adam C, Salomon JC. Immunoglobulins in intact, immunized, and contaminated axenic mice: study of serum IgA. Journal of immunology 1971;107:1647-55. [PubMed] [Google Scholar]
- [20].Ivanov II, Littman DR. Segmented filamentous bacteria take the stage. Mucosal Immunol. 2010;3:209-12. doi: 10.1038/mi.2010.3. PMID:20147894 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [21].Cummings JH, Pomare EW, Branch WJ, Naylor CP, Macfarlane GT. Short chain fatty acids in human large intestine, portal, hepatic and venous blood. Gut 1987;28:1221-7. doi: 10.1136/gut.28.10.1221. PMID:3678950 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [22].Kumar M, Babaei P, Ji B, Nielsen J. Human gut microbiota and healthy aging: Recent developments and future prospective. Nutr Healthy Aging. 2016;4:3-16. doi: 10.3233/NHA-150002. PMID:28035338 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [23].De Filippo C Cavalieri D, Di Paola M, Ramazzotti M, Poullet JB, Massart S, Collini S, Pieraccini G, Lionetti P. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc Natl Acad Sci U S A. 2010;107:14691-6. doi: 10.1073/pnas.1005963107. PMID:20679230 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [24].Frank DN, St Amand AL, Feldman RA, Boedeker EC, Harpaz N, Pace NR. Molecular-phylogenetic characterization of microbial community imbalances in human inflammatory bowel diseases. Proc Natl Acad Sci U S A. 2007;104:13780-5. doi: 10.1073/pnas.0706625104. PMID:17699621 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [25].Martinez-Medina M, Aldeguer X, Gonzalez-Huix F, Acero D, Garcia-Gil LJ. Abnormal microbiota composition in the ileocolonic mucosa of Crohn's disease patients as revealed by polymerase chain reaction-denaturing gradient gel electrophoresis. Inflammatory bowel diseases. 2006;12:1136-45. doi: 10.1097/01.mib.0000235828.09305.0c. PMID:17119388 [DOI] [PubMed] [Google Scholar]
- [26].Arpaia N, Campbell C, Fan X, Dikiy S, van der Veeken J, deRoos P, Liu H, Cross JR, Pfeffer K, Coffer PJ, et al.. Metabolites produced by commensal bacteria promote peripheral regulatory T-cell generation. Nature. 2013;504:451-5. doi: 10.1038/nature12726. PMID:24226773 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [27].Wrzosek L, Miquel S, Noordine ML, Bouet S, Joncquel Chevalier-Curt M, Robert V, Philippe C, Bridonneau C, Cherbuy C, Robbe-Masselot C, et al.. Bacteroides thetaiotaomicron and Faecalibacterium prausnitzii influence the production of mucus glycans and the development of goblet cells in the colonic epithelium of a gnotobiotic model rodent. BMC Biol. 2013;11:61. doi: 10.1186/1741-7007-11-61. PMID:23692866 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [28].Willemsen LE, Koetsier MA, van Deventer SJ, van Tol EA. Short chain fatty acids stimulate epithelial mucin 2 expression through differential effects on prostaglandin E(1) and E(2) production by intestinal myofibroblasts. Gut. 2003;52:1442-7. doi: 10.1136/gut.52.10.1442. PMID:12970137 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [29].Gaudier E, Jarry A, Blottiere HM, de Coppet P, Buisine MP, Aubert JP, Laboisse C, Cherbut C, Hoebler C. Butyrate specifically modulates MUC gene expression in intestinal epithelial goblet cells deprived of glucose. Am J Physiol Gastrointest Liver Physiol. 2004;287:G1168-74. doi: 10.1152/ajpgi.00219.2004. PMID:15308471 [DOI] [PubMed] [Google Scholar]
- [30].Fukuda S, Toh H, Hase K, Oshima K, Nakanishi Y, Yoshimura K, Tobe T, Clarke JM, Topping DL, Suzuki T, et al.. Bifidobacteria can protect from enteropathogenic infection through production of acetate. Nature. 2011;469:543-7. doi: 10.1038/nature09646. PMID:21270894 [DOI] [PubMed] [Google Scholar]
- [31].Peng L, Li ZR, Green RS, Holzman IR, Lin J. Butyrate enhances the intestinal barrier by facilitating tight junction assembly via activation of AMP-activated protein kinase in Caco-2 cell monolayers. The Journal of nutrition. 2009;139:1619-25. doi: 10.3945/jn.109.104638. PMID:19625695 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [32].Valenzano MC, DiGuilio K, Mercado J, Teter M, To J, Ferraro B, Mixson B, Manley I, Baker V, Moore BA, et al.. Remodeling of Tight Junctions and Enhancement of Barrier Integrity of the CACO-2 Intestinal Epithelial Cell Layer by Micronutrients. PloS One. 2015;10:e0133926. doi: 10.1371/journal.pone.0133926. PMID:26226276 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [33].Ivanov II, Atarashi K, Manel N, Brodie EL, Shima T, Karaoz U, Wei D, Goldfarb KC, Santee CA, Lynch SV, et al.. Induction of intestinal Th17 cells by segmented filamentous bacteria. Cell. 2009;139:485-98. doi: 10.1016/j.cell.2009.09.033. PMID:19836068 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [34].Atarashi K, Tanoue T, Shima T, Imaoka A, Kuwahara T, Momose Y, Cheng G, Yamasaki S, Saito T, Ohba Y, et al.. Induction of colonic regulatory T cells by indigenous Clostridium species. Science. 2011;331:337-41. doi: 10.1126/science.1198469. PMID:21205640 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [35].Geuking MB, Cahenzli J, Lawson MA, Ng DC, Slack E, Hapfelmeier S, McCoy KD, Macpherson AJ. Intestinal bacterial colonization induces mutualistic regulatory T cell responses. Immunity. 2011;34:794-806. doi: 10.1016/j.immuni.2011.03.021. PMID:21596591 [DOI] [PubMed] [Google Scholar]
- [36].Round JL, Lee SM, Li J, Tran G, Jabri B, Chatila TA, Mazmanian SK. The Toll-like receptor 2 pathway establishes colonization by a commensal of the human microbiota. Science. 2011;332:974-7. doi: 10.1126/science.1206095. PMID:21512004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [37].Wen L, Ley RE, Volchkov PY, Stranges PB, Avanesyan L, Stonebraker AC, Hu C, Wong FS, Szot GL, Bluestone JA, et al.. Innate immunity and intestinal microbiota in the development of Type 1 diabetes. Nature. 2008;455:1109-13. doi: 10.1038/nature07336. PMID:18806780 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [38].Vaarala O. Is the origin of type 1 diabetes in the gut? Immunol Cell Biol. 2012;90:271-6. doi: 10.1038/icb.2011.115. PMID:22290506 [DOI] [PubMed] [Google Scholar]
- [39].Carstens LE, Westerbeek EA, van Zwol A, van Elburg RM. Neonatal antibiotics in preterm infants and allergic disorders later in life. Pediatr Allergy Immunol. 2016;27:759-64. doi: 10.1111/pai.12614. PMID:27434167 [DOI] [PubMed] [Google Scholar]
- [40].Mulder B, Pouwels KB, Schuiling-Veninga CC, Bos HJ, de Vries TW, Jick SS, Hak E. Antibiotic use during pregnancy and asthma in preschool children: the influence of confounding. Clin Exp Allergy. 2016;46:1214-26. doi: 10.1111/cea.12756. PMID:27159872 [DOI] [PubMed] [Google Scholar]
- [41].Schulfer A, Blaser MJ. Risks of Antibiotic Exposures Early in Life on the Developing Microbiome. PLoS Pathog. 2015;11:e1004903. doi: 10.1371/journal.ppat.1004903. PMID:26135581 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [42].Littman DR, Pamer EG. Role of the commensal microbiota in normal and pathogenic host immune responses. Cell host & microbe. 2011;10:311-23. doi: 10.1016/j.chom.2011.10.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [43].Stefka AT, Feehley T, Tripathi P, Qiu J, McCoy K, Mazmanian SK, Tjota MY, Seo GY, Cao S, Theriault BR, et al.. Commensal bacteria protect against food allergen sensitization. Proc Natl Acad Sci U S A. 2014;111:13145-50. doi: 10.1073/pnas.1412008111. PMID:25157157 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [44].Cox LM, Yamanishi S, Sohn J, Alekseyenko AV, Leung JM, Cho I, Kim SG, Li H, Gao Z, Mahana D, et al.. Altering the intestinal microbiota during a critical developmental window has lasting metabolic consequences. Cell. 2014;158:705-21. doi: 10.1016/j.cell.2014.05.052. PMID:25126780 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [45].Lee YK, Menezes JS, Umesaki Y, Mazmanian SK. Proinflammatory T-cell responses to gut microbiota promote experimental autoimmune encephalomyelitis. Proc Natl Acad Sci U S A. 2011;108(Suppl 1):4615-22. doi: 10.1073/pnas.1000082107. PMID:20660719 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [46].Lee H, Jin BE, Jang E, Lee AR, Han DS, Kim HY, Youn J. Gut-residing Microbes Alter the Host Susceptibility to Autoantibody-mediated Arthritis. Immune Netw. 2014;14:38-44. doi: 10.4110/in.2014.14.1.38. PMID:24605079 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [47].Jin S, Zhao D, Cai C, Song D, Shen J, Xu A, Qiao Y, Ran Z, Zheng Q. Low-dose penicillin exposure in early life decreases Th17 and the susceptibility to DSS colitis in mice through gut microbiota modification. Sci Rep. 2017;7:43662. doi: 10.1038/srep43662. PMID:28272549 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [48].Turnbaugh PJ, Ley RE, Hamady M, Fraser-Liggett CM, Knight R, Gordon JI. The human microbiome project. Nature. 2007;449:804-10. doi: 10.1038/nature06244. PMID:17943116 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [49].Sudo N, Sawamura S, Tanaka K, Aiba Y, Kubo C, Koga Y. The requirement of intestinal bacterial flora for the development of an IgE production system fully susceptible to oral tolerance induction. Journal of immunology 1997;159:1739-45. [PubMed] [Google Scholar]
- [50].Qin J, Li R, Raes J, Arumugam M, Burgdorf KS, Manichanh C, Nielsen T, Pons N, Levenez F, Yamada T, et al.. A human gut microbial gene catalogue established by metagenomic sequencing. Nature. 2010;464:59-65. doi: 10.1038/nature08821. PMID:20203603 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [51].Rodriguez JM, Murphy K, Stanton C, Ross RP, Kober OI, Juge N, Avershina E, Rudi K, Narbad A, Jenmalm MC, et al.. The composition of the gut microbiota throughout life, with an emphasis on early life. Microb Ecol Health Dis. 2015;26:26050. PMID:25651996 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [52].Ramanan D, Tang MS, Bowcutt R, Loke P, Cadwell K. Bacterial sensor Nod2 prevents inflammation of the small intestine by restricting the expansion of the commensal Bacteroides vulgatus. Immunity. 2014;41:311-24. doi: 10.1016/j.immuni.2014.06.015. PMID:25088769 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [53].Tamburini S, Shen N, Wu HC, Clemente JC. The microbiome in early life: implications for health outcomes. Nat Med. 2016;22:713-22. doi: 10.1038/nm.4142. PMID:27387886 [DOI] [PubMed] [Google Scholar]
- [54].Wegmann TG, Lin H, Guilbert L, Mosmann TR. Bidirectional cytokine interactions in the maternal-fetal relationship: is successful pregnancy a TH2 phenomenon? Immunol Today 1993;14:353-6. doi: 10.1016/0167-5699(93)90235-D. PMID:8363725 [DOI] [PubMed] [Google Scholar]
- [55].Marzi M, Vigano A, Trabattoni D, Villa ML, Salvaggio A, Clerici E, Clerici M. Characterization of type 1 and type 2 cytokine production profile in physiologic and pathologic human pregnancy. Clin Exp Immunol. 1996;106:127-33. doi: 10.1046/j.1365-2249.1996.d01-809.x. PMID:8870710 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [56].Tsuda H, Michimata T, Hayakawa S, Tanebe K, Sakai M, Fujimura M, Matsushima K, Saito S. A Th2 chemokine, TARC, produced by trophoblasts and endometrial gland cells, regulates the infiltration of CCR4+ T lymphocytes into human decidua at early pregnancy. Am J Reprod Immunol. 2002;48:1-8. doi: 10.1034/j.1600-0897.2002.01117.x. PMID:12322891 [DOI] [PubMed] [Google Scholar]
- [57].McFadden JP, Thyssen JP, Basketter DA, Puangpet P, Kimber I. T helper cell 2 immune skewing in pregnancy/early life: chemical exposure and the development of atopic disease and allergy. Br J Dermatol. 2015;172:584-91. doi: 10.1111/bjd.13497. PMID:25354210 [DOI] [PubMed] [Google Scholar]
- [58].Riedler J, Braun-Fahrlander C, Eder W, Schreuer M, Waser M, Maisch S, Carr D, Schierl R, Nowak D, von Mutius E, et al.. Exposure to farming in early life and development of asthma and allergy: a cross-sectional survey. Lancet. 2001;358:1129-33. doi: 10.1016/S0140-6736(01)06252-3. PMID:11597666 [DOI] [PubMed] [Google Scholar]
- [59].Kopp MV, Zehle C, Pichler J, Szepfalusi Z, Moseler M, Deichmann K, Forster J, Kuehr J. Allergen-specific T cell reactivity in cord blood: the influence of maternal cytokine production. Clin Exp Allergy. 2001;31:1536-43. doi: 10.1046/j.1365-2222.2001.01198.x. PMID:11678853 [DOI] [PubMed] [Google Scholar]
- [60].Lehmann I, Thoelke A, Rehwagen M, Rolle-Kampczyk U, Schlink U, Schulz R, Borte M, Diez U, Herbarth O. The influence of maternal exposure to volatile organic compounds on the cytokine secretion profile of neonatal T cells. Environ Toxicol. 2002;17:203-10. doi: 10.1002/tox.10055. PMID:12112628 [DOI] [PubMed] [Google Scholar]
- [61].Satokari R, Gronroos T, Laitinen K, Salminen S, Isolauri E. Bifidobacterium and Lactobacillus DNA in the human placenta. Lett Appl Microbiol. 2009;48:8-12. doi: 10.1111/j.1472-765X.2008.02475.x. PMID:19018955 [DOI] [PubMed] [Google Scholar]
- [62].Aagaard K, Ma J, Antony KM, Ganu R, Petrosino J, Versalovic J. The placenta harbors a unique microbiome. Sci Transl Med. 2014;6:237ra65. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [63].Steel JH, O'Donoghue K, Kennea NL, Sullivan MH, Edwards AD. Maternal origin of inflammatory leukocytes in preterm fetal membranes, shown by fluorescence in situ hybridisation. Placenta. 2005;26:672-7. doi: 10.1016/j.placenta.2004.10.003. PMID:16085046 [DOI] [PubMed] [Google Scholar]
- [64].Jimenez E, Fernandez L, Marin ML, Martin R, Odriozola JM, Nueno-Palop C, Narbad A, Olivares M, Xaus J, Rodríguez JM. Isolation of commensal bacteria from umbilical cord blood of healthy neonates born by cesarean section. Curr Microbiol. 2005;51:270-4. doi: 10.1007/s00284-005-0020-3. PMID:16187156 [DOI] [PubMed] [Google Scholar]
- [65].Jimenez E, Marin ML, Martin R, Odriozola JM, Olivares M, Xaus J, Fernández L, Rodríguez JM. Is meconium from healthy newborns actually sterile? Res Microbiol. 2008;159:187-93. doi: 10.1016/j.resmic.2007.12.007. PMID:18281199 [DOI] [PubMed] [Google Scholar]
- [66].Ardissone AN, de la Cruz DM, Davis-Richardson AG, Rechcigl KT, Li N, Drew JC, Murgas-Torrazza R, Sharma R, Hudak ML, Triplett EW, et al.. Meconium microbiome analysis identifies bacteria correlated with premature birth. PloS One. 2014;9:e90784. doi: 10.1371/journal.pone.0090784. PMID:24614698 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [67].Tojo R, Suarez A, Clemente MG, de los Reyes-Gavilan CG, Margolles A, Gueimonde M, Ruas-Madiedo P. Intestinal microbiota in health and disease: role of bifidobacteria in gut homeostasis. World J Gastroenterol. 2014;20:15163-76. doi: 10.3748/wjg.v20.i41.15163. PMID:25386066 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [68].Gerhold K, Avagyan A, Seib C, Frei R, Steinle J, Ahrens B, Dittrich AM, Blumchen K, Lauener R, Hamelmann E. Prenatal initiation of endotoxin airway exposure prevents subsequent allergen-induced sensitization and airway inflammation in mice. J Allergy Clin Immunol. 2006;118:666-73. doi: 10.1016/j.jaci.2006.05.022. PMID:16950286 [DOI] [PubMed] [Google Scholar]
- [69].Blumer N, Herz U, Wegmann M, Renz H. Prenatal lipopolysaccharide-exposure prevents allergic sensitization and airway inflammation, but not airway responsiveness in a murine model of experimental asthma. Clin Exp Allergy. 2005;35:397-402. doi: 10.1111/j.1365-2222.2005.02184.x. PMID:15784121 [DOI] [PubMed] [Google Scholar]
- [70].Conrad ML, Ferstl R, Teich R, Brand S, Blumer N, Yildirim AO, Patrascan CC, Hanuszkiewicz A, Akira S, Wagner H, et al.. Maternal TLR signaling is required for prenatal asthma protection by the nonpathogenic microbe Acinetobacter lwoffii F78. J Exp Med. 2009;206:2869-77. doi: 10.1084/jem.20090845. PMID:19995952 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [71].Dominguez-Bello MG, Costello EK, Contreras M, Magris M, Hidalgo G, Fierer N, Knight R. Delivery mode shapes the acquisition and structure of the initial microbiota across multiple body habitats in newborns. Proc Natl Acad Sci U S A. 2010;107:11971-5. doi: 10.1073/pnas.1002601107. PMID:20566857 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [72].Kristensen K, Henriksen L. Cesarean section and disease associated with immune function. J Allergy Clin Immunol. 2016;137:587-90. doi: 10.1016/j.jaci.2015.07.040. PMID:26371844 [DOI] [PubMed] [Google Scholar]
- [73].Lotz M, Gutle D, Walther S, Menard S, Bogdan C, Hornef MW. Postnatal acquisition of endotoxin tolerance in intestinal epithelial cells. J Exp Med. 2006;203:973-84. doi: 10.1084/jem.20050625. PMID:16606665 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [74].Brandtzaeg P. The mucosal immune system and its integration with the mammary glands. J Ped. 2010;156:S8-15. doi: 10.1016/j.jpeds.2009.11.014. [DOI] [PubMed] [Google Scholar]
- [75].Lonnerdal B. Infant formula and infant nutrition: bioactive proteins of human milk and implications for composition of infant formulas. Am J Clin Nutr. 2014;99:712S-7S. doi: 10.3945/ajcn.113.071993. PMID:24452231 [DOI] [PubMed] [Google Scholar]
- [76].Fichter M, Klotz M, Hirschberg DL, Waldura B, Schofer O, Ehnert S, Schwarz LK, Ginneken CV, Schäfer KH. Breast milk contains relevant neurotrophic factors and cytokines for enteric nervous system development. Mol Nutr Food Res. 2011;55:1592-6. doi: 10.1002/mnfr.201100124. PMID:21809438 [DOI] [PubMed] [Google Scholar]
- [77].Fernandez L, Langa S, Martin V, Jimenez E, Martin R, Rodriguez JM. The microbiota of human milk in healthy women. Cell Mol Biol (Noisy-le-grand). 2013;59:31-42. PMID:24200019 [PubMed] [Google Scholar]
- [78].McGuire W, Anthony MY. Donor human milk versus formula for preventing necrotising enterocolitis in preterm infants: systematic review. Arch Dis Child Fetal Neonatal Ed. 2003;88:F11-4. doi: 10.1136/fn.88.1.F11. PMID:12496220 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [79].Stuebe A. The risks of not breastfeeding for mothers and infants. Rev Obstet Gynecol. 2009;2:222-31. PMID:20111658 [PMC free article] [PubMed] [Google Scholar]
- [80].Cani PD, Delzenne NM. The role of the gut microbiota in energy metabolism and metabolic disease. Curr Pharm Des. 2009;15:1546-58. doi: 10.2174/138161209788168164. PMID:19442172 [DOI] [PubMed] [Google Scholar]
- [81].Yu LC, Wang JT, Wei SC, Ni YH. Host-microbial interactions and regulation of intestinal epithelial barrier function: From physiology to pathology. World journal of gastrointestinal pathophysiology. 2012;3:27-43. doi: 10.4291/wjgp.v3.i1.27. PMID:22368784 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [82].Ge X, Ding C, Zhao W, Xu L, Tian H, Gong J, Zhu M, Li J, Li N. Antibiotics-induced depletion of mice microbiota induces changes in host serotonin biosynthesis and intestinal motility. J Transl Med. 2017;15:13. doi: 10.1186/s12967-016-1105-4. PMID:28086815 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [83].Abrams GD, Bishop JE. Effect of the normal microbial flora on gastrointestinal motility. Proc Soc Exp Biol Med 1967;126:301-4. doi: 10.3181/00379727-126-32430. PMID:6066182 [DOI] [PubMed] [Google Scholar]
- [84].Husebye E, Hellstrom PM, Sundler F, Chen J, Midtvedt T. Influence of microbial species on small intestinal myoelectric activity and transit in germ-free rats. Am J Physiol Gastrointest Liver Physiol. 2001;280:G368-80. PMID:11171619 [DOI] [PubMed] [Google Scholar]
- [85].Wiles TJ, Jemielita M, Baker RP, Schlomann BH, Logan SL, Ganz J, Melancon E, Eisen JS, Guillemin K, Parthasarathy R. Host Gut Motility Promotes Competitive Exclusion within a Model Intestinal Microbiota. PLoS Biol. 2016;14:e1002517. doi: 10.1371/journal.pbio.1002517. PMID:27458727 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [86].Rolig AS, Mittge EK, Ganz J, Troll JV, Melancon E, Wiles TJ, Alligood K, Stephens WZ, Eisen JS, Guillemin K. The enteric nervous system promotes intestinal health by constraining microbiota composition. PLoS Biol. 2017;15:e2000689. doi: 10.1371/journal.pbio.2000689. PMID:28207737 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [87].Johansson ME, Gustafsson JK, Holmen-Larsson J, Jabbar KS, Xia L, Xu H, Ghishan FK, Carvalho FA, Gewirtz AT, Sjövall H, et al.. Bacteria penetrate the normally impenetrable inner colon mucus layer in both murine colitis models and patients with ulcerative colitis. Gut. 2014;63:281-91. doi: 10.1136/gutjnl-2012-303207. PMID:23426893 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [88].Johansson ME, Jakobsson HE, Holmen-Larsson J, Schutte A, Ermund A, Rodriguez-Pineiro AM, Arike L, Wising C, Svensson F, Bäckhed F, et al.. Normalization of Host Intestinal Mucus Layers Requires Long-Term Microbial Colonization. Cell Host Microbe. 2015;18:582-92. doi: 10.1016/j.chom.2015.10.007. PMID:26526499 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [89].Jakobsson HE, Rodriguez-Pineiro AM, Schutte A, Ermund A, Boysen P, Bemark M, Sommer F, Bäckhed F, Hansson GC, Johansson ME. The composition of the gut microbiota shapes the colon mucus barrier. EMBO Rep. 2015;16:164-77. doi: 10.15252/embr.201439263. PMID:25525071 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [90].Gallagher P, O'Mahony D. Constipation in old age. Best Pract Res Clin Gastroenterol. 2009;23:875-87. doi: 10.1016/j.bpg.2009.09.001. PMID:19942165 [DOI] [PubMed] [Google Scholar]
- [91].Saffrey MJ. Aging of the mammalian gastrointestinal tract: A complex organ system. Age (Dordr). 2014;36:9603. doi: 10.1007/s11357-013-9603-2. PMID:24352567 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [92].Hoyle CH, Saffrey MJ. Effects of aging on cholinergic neuromuscular transmission in isolated small intestine of ad libitum fed and calorically-restricted rats. Neurogastroenterol Motil. 2012;24:586-92. doi: 10.1111/j.1365-2982.2012.01913.x. PMID:22435850 [DOI] [PubMed] [Google Scholar]
- [93].Bitar KN, Patil SB. Aging and gastrointestinal smooth muscle. Mech Ageing Dev. 2004;125:907-10. doi: 10.1016/j.mad.2004.05.010. PMID:15563937 [DOI] [PubMed] [Google Scholar]
- [94].Lopes GS, Mora OA, Cerri P, Faria FP, Jurkiewicz NH, Jurkiewicz A, Smaili SS. Mitochondrial alterations and apoptosis in smooth muscle from aged rats. Biochim Biophys Acta. 2004;1658:187-94. doi: 10.1016/j.bbabio.2004.05.011. PMID:15450956 [DOI] [PubMed] [Google Scholar]
- [95].Saffrey MJ. Cellular changes in the enteric nervous system during ageing. Dev Biol. 2013;382:344-55. doi: 10.1016/j.ydbio.2013.03.015. PMID:23537898 [DOI] [PubMed] [Google Scholar]
- [96].Phillips RJ, Kieffer EJ, Powley TL. Aging of the myenteric plexus: neuronal loss is specific to cholinergic neurons. Auton Neurosci. 2003;106:69-83. doi: 10.1016/S1566-0702(03)00072-9. PMID:12878075 [DOI] [PubMed] [Google Scholar]
- [97].Feher E, Penzes L. Density of substance P, vasoactive intestinal polypeptide and somatostatin-containing nerve fibers in the ageing small intestine of the rats. Gerontology 1987;33:341-8. doi: 10.1159/000212901. PMID:2450811 [DOI] [PubMed] [Google Scholar]
- [98].Corns RA, Hidaka H, Santer RM. Neurocalcin-alpha immunoreactivity in the enteric nervous system of young and aged rats. Cell Calcium. 2002;31:53-8. doi: 10.1054/ceca.2001.0261. PMID:11969245 [DOI] [PubMed] [Google Scholar]
- [99].Kirkwood TB. Intrinsic ageing of gut epithelial stem cells. Mech Ageing Dev. 2004;125:911-5. doi: 10.1016/j.mad.2004.09.004. PMID:15563938 [DOI] [PubMed] [Google Scholar]
- [100].Jurk D, Wang C, Miwa S, Maddick M, Korolchuk V, Tsolou A, Gonos ES, Thrasivoulou C, Saffrey MJ, Cameron K, et al.. Postmitotic neurons develop a p21-dependent senescence-like phenotype driven by a DNA damage response. Aging Cell. 2012;11:996-1004. doi: 10.1111/j.1474-9726.2012.00870.x. PMID:22882466 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [101].Ershler WB. Biological interactions of aging and anemia: A focus on cytokines. J Am Geriatr Soc. 2003;51:S18-21. doi: 10.1046/j.1532-5415.51.3s.2.x. PMID:12588568 [DOI] [PubMed] [Google Scholar]
- [102].Rimoldi M, Chieppa M, Salucci V, Avogadri F, Sonzogni A, Sampietro GM, Nespoli A, Viale G, Allavena P, Rescigno M. Intestinal immune homeostasis is regulated by the crosstalk between epithelial cells and dendritic cells. Nat Immunol. 2005;6:507-14. doi: 10.1038/ni1192. PMID:15821737 [DOI] [PubMed] [Google Scholar]
- [103].Spadoni I, Iliev ID, Rossi G, Rescigno M. Dendritic cells produce TSLP that limits the differentiation of Th17 cells, fosters Treg development, and protects against colitis. Mucosal Immunol. 2012;5:184-93. doi: 10.1038/mi.2011.64. PMID:22236997 [DOI] [PubMed] [Google Scholar]
- [104].Thrasivoulou C, Soubeyre V, Ridha H, Giuliani D, Giaroni C, Michael GJ, Saffrey MJ, Cowen T. Reactive oxygen species, dietary restriction and neurotrophic factors in age-related loss of myenteric neurons. Aging Cell. 2006;5:247-57. doi: 10.1111/j.1474-9726.2006.00214.x. PMID:16842497 [DOI] [PubMed] [Google Scholar]
- [105].Schiffrin EJ, Morley JE, Donnet-Hughes A, Guigoz Y. The inflammatory status of the elderly: the intestinal contribution. Mutat Res. 2010;690:50-6. doi: 10.1016/j.mrfmmm.2009.07.011. PMID:19666034 [DOI] [PubMed] [Google Scholar]
- [106].Biagi E, Nylund L, Candela M, Ostan R, Bucci L, Pini E, Nikkïla J, Monti D, Satokari R, Franceschi C, et al.. Through ageing, and beyond: gut microbiota and inflammatory status in seniors and centenarians. PloS One. 2010;5:e10667. doi: 10.1371/journal.pone.0010667. PMID:20498852 [DOI] [PMC free article] [PubMed] [Google Scholar]