

Multivalent structure of an alphabetaT cell receptor
- PMID: 9990061
- PMCID: PMC15512
- DOI: 10.1073/pnas.96.4.1547
Multivalent structure of an alphabetaT cell receptor
Abstract
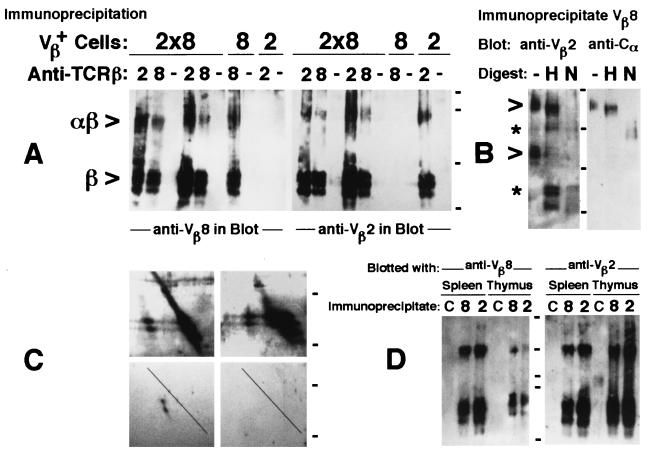
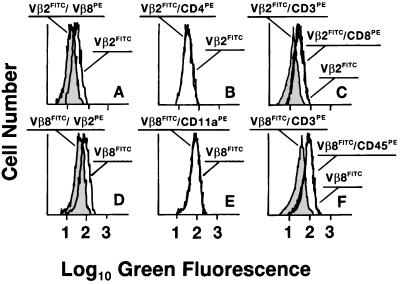
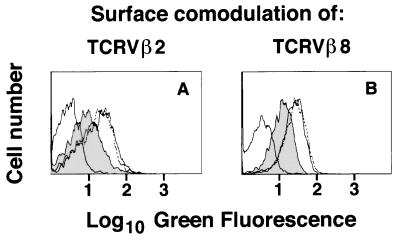
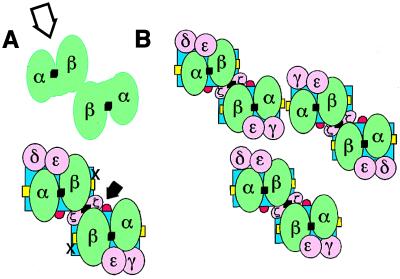
Whether there is one or multiple alphabetaT cell antigen receptor (TCR) recognition modules in a given TCR/CD3 complex is a long-standing controversy in immunology. We show that T cells from transgenic mice that coexpress comparable amounts of two distinct TCRbeta chains incorporate at least two alphabetaTCRs in a single TCR/CD3 complex. Evidence for bispecific alphabetaTCRs was obtained by immunoprecipitation and immunoblotting and confirmed on the surface of living cells both by fluorescence resonance energy transfer and comodulation assays by using antibodies specific for TCRbeta-variable regions. Such (alphabeta)2TCR/CD3 or higher-order complexes were evident in T cells studied either ex vivo or after expansion in vitro. T cell activation is thought by many, but not all, to require TCR cross-linking by its antigen/major histocompatibility complex ligand. The implications of a multivalent (alphabeta)2TCR/CD3 complex stoichiometry for the ordered docking of specific antigen/major histocompatibility complex, CD4, or CD8 coreceptors and additional TCRs are discussed.
Figures
Similar articles
-
Stoichiometry of the T cell antigen receptor (TCR) complex: each TCR/CD3 complex contains one TCR alpha, one TCR beta, and two CD3 epsilon chains.J Exp Med. 1994 Aug 1;180(2):587-93. doi: 10.1084/jem.180.2.587. J Exp Med. 1994. PMID: 8046335 Free PMC article.
-
Regulation of T cell receptor (TCR) beta gene expression by CD3 complex signaling in immature thymocytes: implications for TCRbeta allelic exclusion.Proc Natl Acad Sci U S A. 1999 Mar 30;96(7):3882-7. doi: 10.1073/pnas.96.7.3882. Proc Natl Acad Sci U S A. 1999. PMID: 10097132 Free PMC article.
-
Early TCR alpha beta expression promotes maturation of T cells expressing Fc epsilon RI gamma containing TCR/CD3 complexes.J Immunol. 2001 Jun 1;166(11):6616-24. doi: 10.4049/jimmunol.166.11.6616. J Immunol. 2001. PMID: 11359815
-
αβ and γδ T cell receptors: Similar but different.J Leukoc Biol. 2020 Jun;107(6):1045-1055. doi: 10.1002/JLB.2MR1219-233R. Epub 2020 Jan 29. J Leukoc Biol. 2020. PMID: 31994778 Review.
-
Thymic selection and tolerance.Transplant Proc. 1996 Dec;28(6):3429-30. Transplant Proc. 1996. PMID: 8962337 Review. No abstract available.
Cited by
-
Early T-cell activation biophysics.HFSP J. 2009 Dec;3(6):401-11. doi: 10.2976/1.3254098. Epub 2009 Nov 10. HFSP J. 2009. PMID: 20514131 Free PMC article.
-
Molecular mechanisms for the assembly of the T cell receptor-CD3 complex.Mol Immunol. 2004 Apr;40(18):1295-305. doi: 10.1016/j.molimm.2003.11.017. Mol Immunol. 2004. PMID: 15072848 Free PMC article. Review.
-
TCR Nanoclusters as the Framework for Transmission of Conformational Changes and Cooperativity.Front Immunol. 2012 May 9;3:115. doi: 10.3389/fimmu.2012.00115. eCollection 2012. Front Immunol. 2012. PMID: 22582078 Free PMC article.
-
Structure of a covalently stabilized complex of a human alphabeta T-cell receptor, influenza HA peptide and MHC class II molecule, HLA-DR1.EMBO J. 2000 Nov 1;19(21):5611-24. doi: 10.1093/emboj/19.21.5611. EMBO J. 2000. PMID: 11060013 Free PMC article.
-
A variable immunoreceptor in a subpopulation of human neutrophils.Proc Natl Acad Sci U S A. 2006 Sep 26;103(39):14441-6. doi: 10.1073/pnas.0603406103. Epub 2006 Sep 18. Proc Natl Acad Sci U S A. 2006. PMID: 16983085 Free PMC article.
References
-
- Meuer S C, Acuto O, Hussey R E, Hodgdon J C, Fitzgerald K A, Schlossman S F, Reinherz E L. Nature (London) 1983;303:808–810. - PubMed
-
- Clevers H, Alarcon B, Wileman T, Terhorst C. Annu Rev Immunol. 1988;6:629–662. - PubMed
-
- Klausner R D, Lippcott-Schwartz J, Bonifacino J S. Annu Rev Immunol. 1990;6:403–431. - PubMed
-
- Garcia K C, Degano M, Stanfield R L, Brunmark A, Jackson M R, Peterson P A, Teyton L, Wilson I A. Science. 1996;274:209–219. - PubMed
-
- Garboczi D N, Ghosh P, Utz U, Fan Q R, Biddison W E, Wiley D C. Nature (London) 1996;384:134–141. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
