

Modification of activity-dependent increases of cerebral blood flow by excitatory synaptic activity and spikes in rat cerebellar cortex
- PMID: 9763643
- PMCID: PMC2231204
- DOI: 10.1111/j.1469-7793.1998.555be.x
Modification of activity-dependent increases of cerebral blood flow by excitatory synaptic activity and spikes in rat cerebellar cortex
Abstract
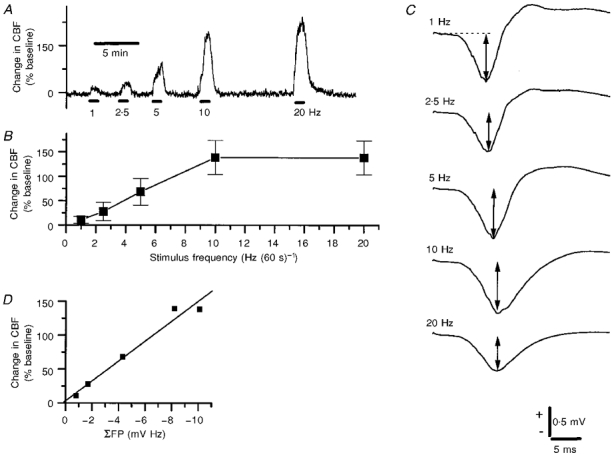
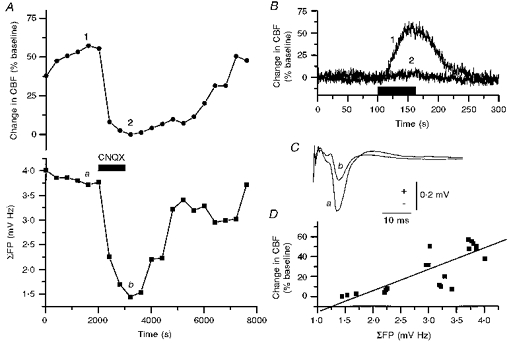
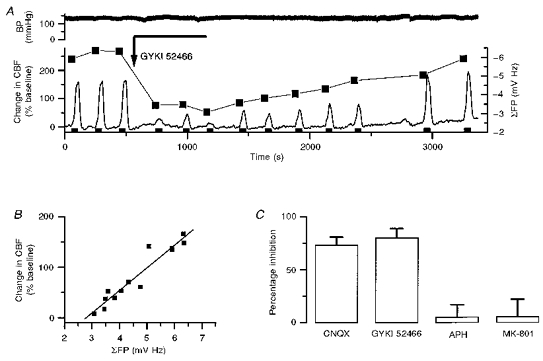
1. Mechanisms of activity-dependent increases in cerebral blood flow (CBF) were examined in rat cerebellar cortex using the laser Doppler flow technique and extracellular recordings of single unit activity and field potentials. 2. Stimulation of the monosynaptic climbing fibre system evoked long-lasting complex spikes in Purkinje cells, and extracellular field potentials with a characteristic profile that indicated contributions from both passive and active membrane mechanisms. The concomitant CBF increases were reproducible at fairly short intervals, and suggest that both synaptic activity and spikes may contribute to increased CBF. 3. Stimulation of the disynaptic parallel fibre system inhibited the spiking activity in Purkinje cells, while the postsynaptic activity increased as indicated by the simultaneously recorded field potential. Nevertheless, CBF always increased. The inhibition of spike firing activity was partly dependent on GABAergic transmission, but may also relate to the intrinsic membrane properties of Purkinje cells. 4. The CBF increases evoked by parallel or climbing fibre stimulation were highly correlated to the sum of neural activities, i.e. the negativity of field potentials multiplied by the stimulus frequency. This suggests a robust link between extracellular current flow and activity-dependent increases in CBF. 5. AMPA receptor blockade attenuated CBF increases and field potential amplitudes, while NMDA receptor antagonism did not. This is consistent with the idea that the CBF responses are of neuronal origin. 6. This study has shown that activity-dependent CBF increases evoked by stimulation of cerebellar parallel fibres are dependent on synaptic excitation, including excitation of inhibitory interneurones, whereas the net activity of Purkinje cells, the principal neurones of the cerebellar cortex, is unimportant for the vascular response. For the climbing fibre system, not only synaptic activity but also the generation of complex spikes from Purkinje cells contribute to the increases in CBF. The strong correlation between CBF and field potential amplitudes suggests that extracellular ion fluxes contribute to the coupling of brain activity to blood flow.
Figures
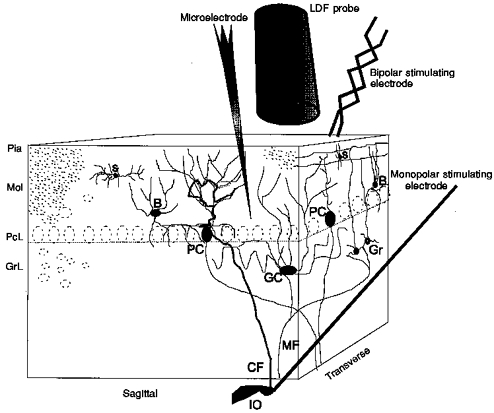
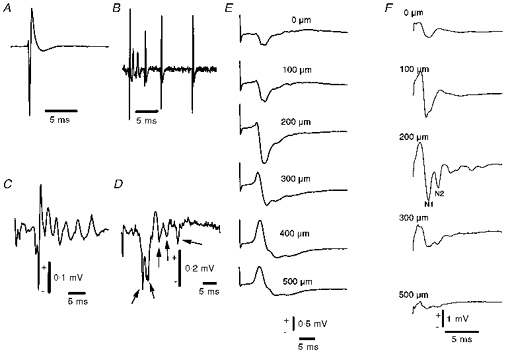
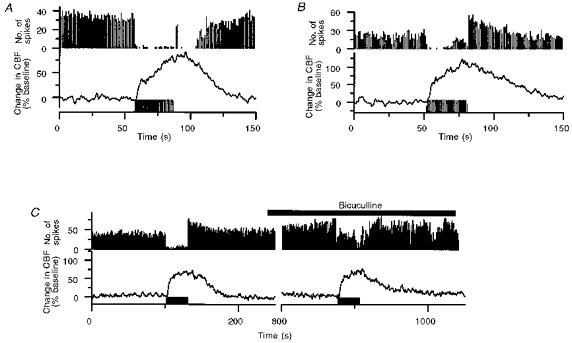
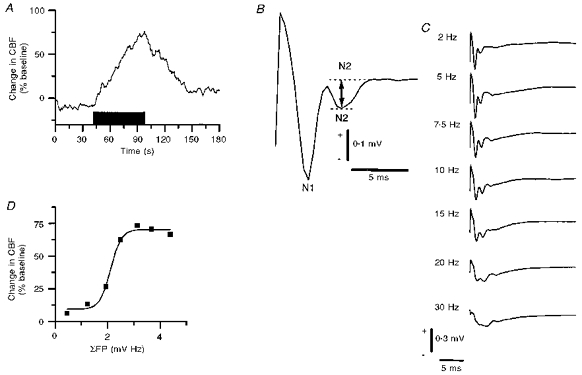
Similar articles
-
Laminar analysis of activity-dependent increases of CBF in rat cerebellar cortex: dependence on synaptic strength.Am J Physiol. 1997 Sep;273(3 Pt 2):H1166-76. doi: 10.1152/ajpheart.1997.273.3.H1166. Am J Physiol. 1997. PMID: 9321803
-
Cerebral blood flow increases evoked by electrical stimulation of rat cerebellar cortex: relation to excitatory synaptic activity and nitric oxide synthesis.Brain Res. 1996 Feb 26;710(1-2):204-14. doi: 10.1016/0006-8993(95)01354-7. Brain Res. 1996. PMID: 8963660
-
Dissociation of spikes, synaptic activity, and activity-dependent increments in rat cerebellar blood flow by tonic synaptic inhibition.Proc Natl Acad Sci U S A. 2003 Dec 23;100(26):16000-5. doi: 10.1073/pnas.2635195100. Epub 2003 Dec 12. Proc Natl Acad Sci U S A. 2003. PMID: 14673091 Free PMC article.
-
The organization of cerebellar cortical circuitry revisited: implications for function.Ann N Y Acad Sci. 2002 Dec;978:135-55. doi: 10.1111/j.1749-6632.2002.tb07562.x. Ann N Y Acad Sci. 2002. PMID: 12582048 Review.
-
Modelling the cerebellar Purkinje cell: experiments in computo.Prog Brain Res. 1994;102:427-41. doi: 10.1016/s0079-6123(08)60557-9. Prog Brain Res. 1994. PMID: 7800831 Review.
Cited by
-
The DIVA model: A neural theory of speech acquisition and production.Lang Cogn Process. 2011 Jan 1;26(7):952-981. doi: 10.1080/01690960903498424. Lang Cogn Process. 2011. PMID: 23667281 Free PMC article.
-
An approach to probe some neural systems interaction by functional MRI at neural time scale down to milliseconds.Proc Natl Acad Sci U S A. 2000 Sep 26;97(20):11026-31. doi: 10.1073/pnas.97.20.11026. Proc Natl Acad Sci U S A. 2000. PMID: 11005873 Free PMC article.
-
Activation of human medial prefrontal cortex during autonomic responses to hypoglycemia.Proc Natl Acad Sci U S A. 2004 Apr 20;101(16):6217-21. doi: 10.1073/pnas.0307048101. Epub 2004 Mar 16. Proc Natl Acad Sci U S A. 2004. PMID: 15026569 Free PMC article.
-
Complex relationship between BOLD signal and synchronization/desynchronization of human brain MEG oscillations.Hum Brain Mapp. 2007 Sep;28(9):805-16. doi: 10.1002/hbm.20322. Hum Brain Mapp. 2007. PMID: 17133396 Free PMC article.
-
Early adaptations in somatosensory cortex after focal ischemic injury to motor cortex.Exp Brain Res. 2006 Jan;168(1-2):178-85. doi: 10.1007/s00221-005-0077-z. Epub 2005 Jul 23. Exp Brain Res. 2006. PMID: 16041499
References
-
- Akgören N, Dalgaard P, Lauritzen M. Cerebral blood flow increases evoked by electrical stimulation of rat cerebellar cortex: relation to excitatory synaptic activity and nitric oxide synthesis. Brain Research. 1996;710:204–214. 10.1016/0006-8993(95)01354-7. - DOI - PubMed
-
- Akgören N, Mathiesen C, Rubin I, Lauritzen M. Laminar analysis of activity-dependent increases of CBF in rat cerebellar cortex: dependence on synaptic strength. American Journal of Physiology. 1997;273:H1166–1176. - PubMed
-
- Andersen P, Eccles JC, Voorhoeve PE. Postsynaptic inhibition of cerebellar Purkinje cells. Journal of Neurophysiology. 1965;27:1138–1153. - PubMed
-
- Azizi SA, Woodward DJ. Inferior olivary nuclear complex of the rat: Morphology and comments on the principles of organization within the olivocerebellar system. Journal of Comparative Neurology. 1987;263:467–484. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources