

Annexin XIIIb associates with lipid microdomains to function in apical delivery
- PMID: 9744874
- PMCID: PMC2141766
- DOI: 10.1083/jcb.142.6.1413
Annexin XIIIb associates with lipid microdomains to function in apical delivery
Abstract
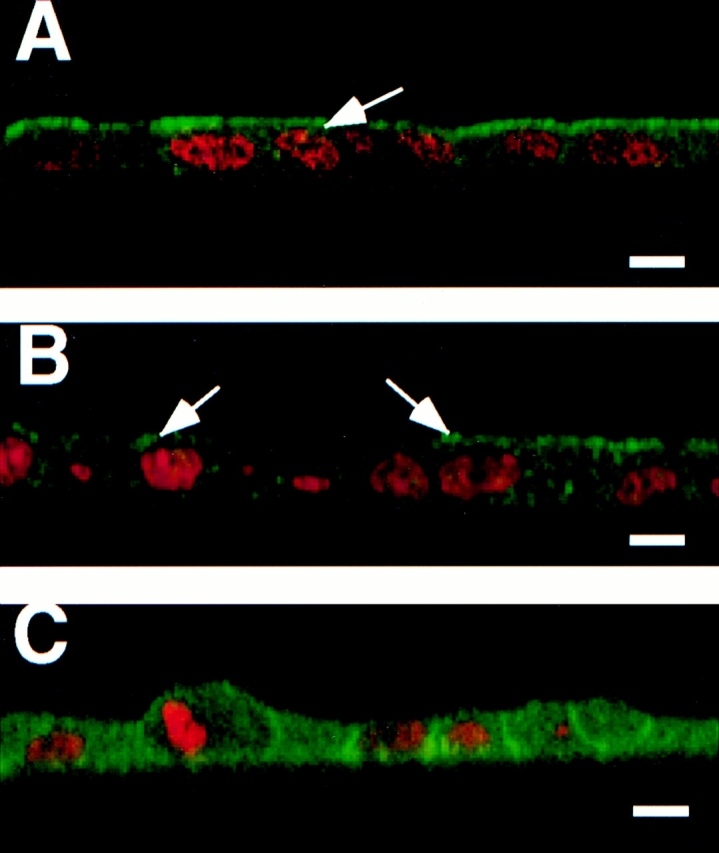
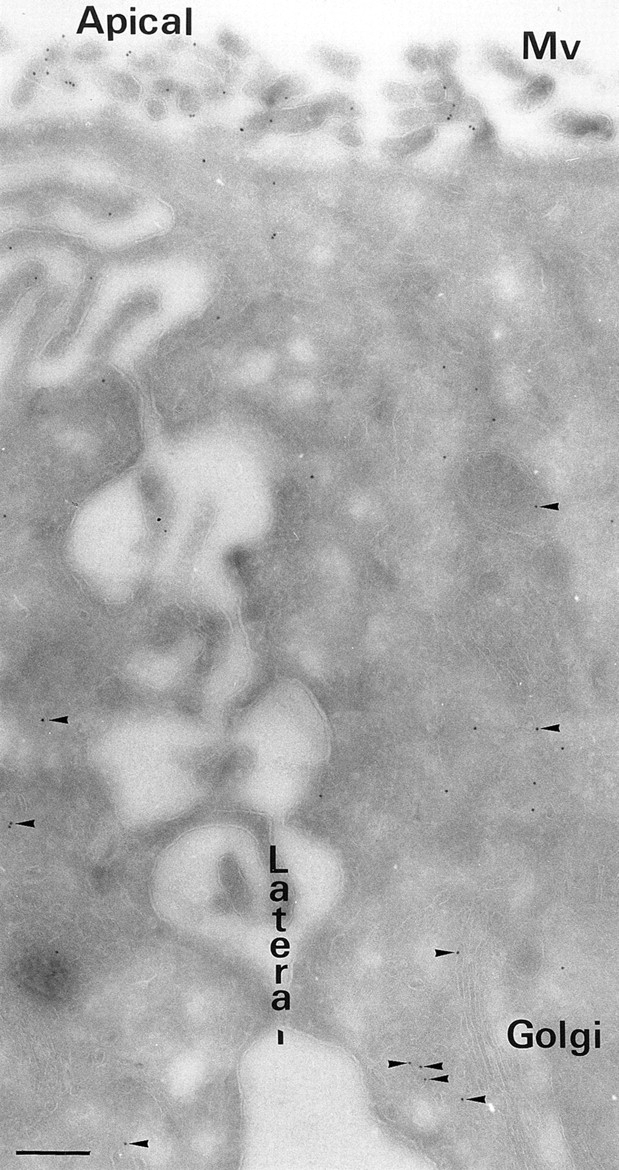
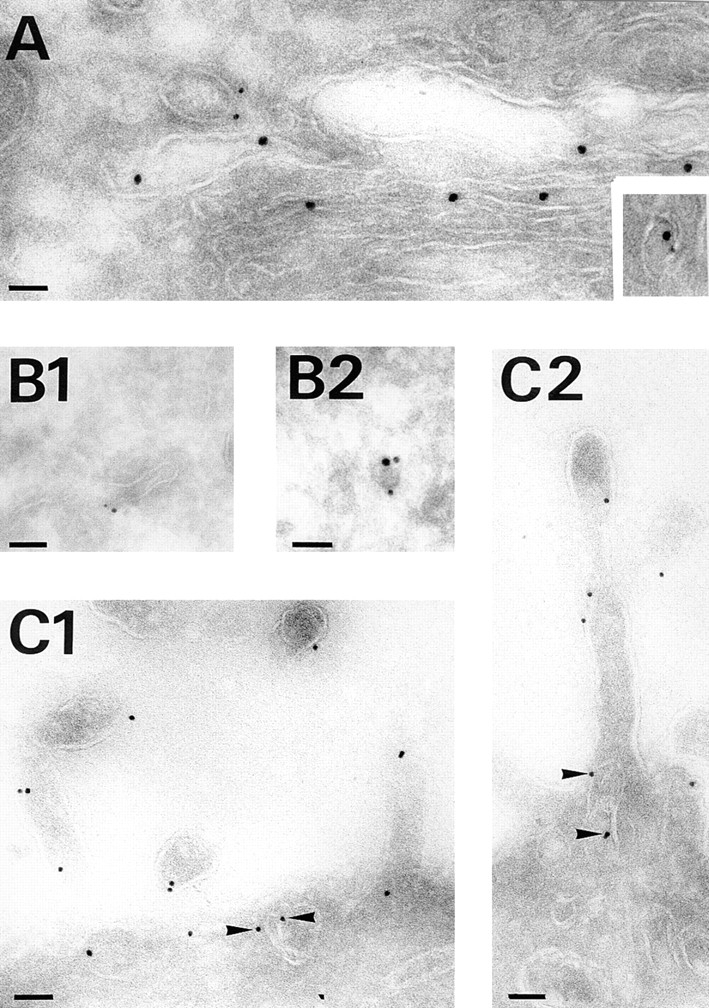
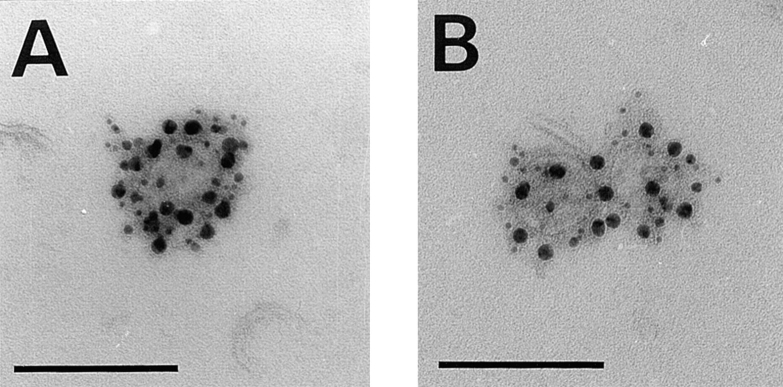
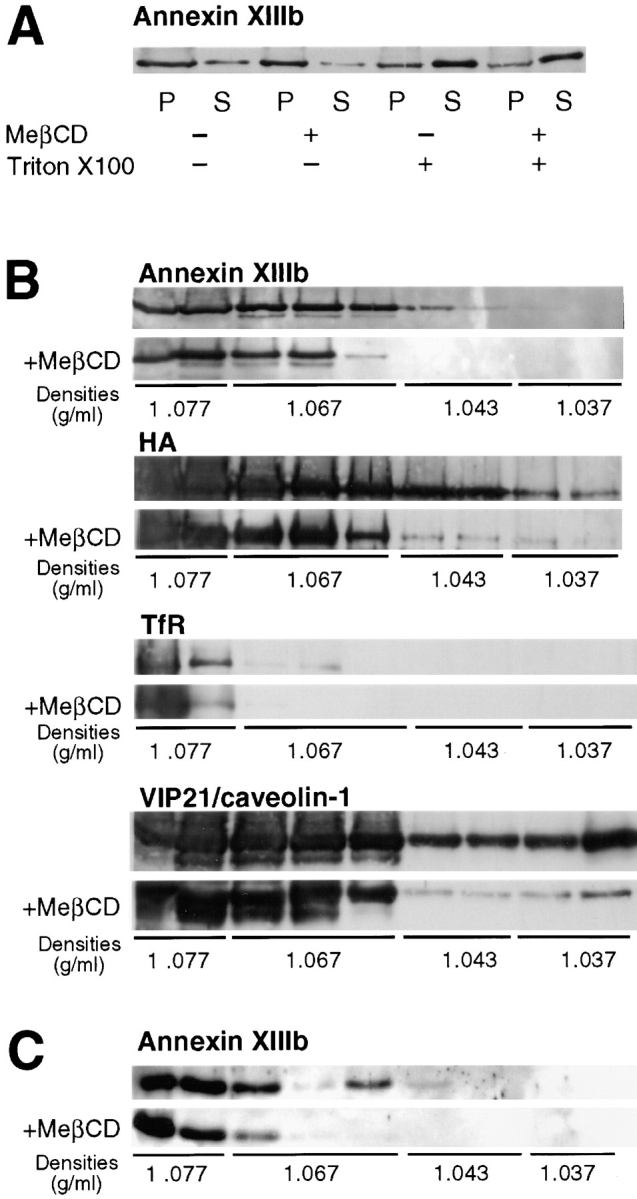
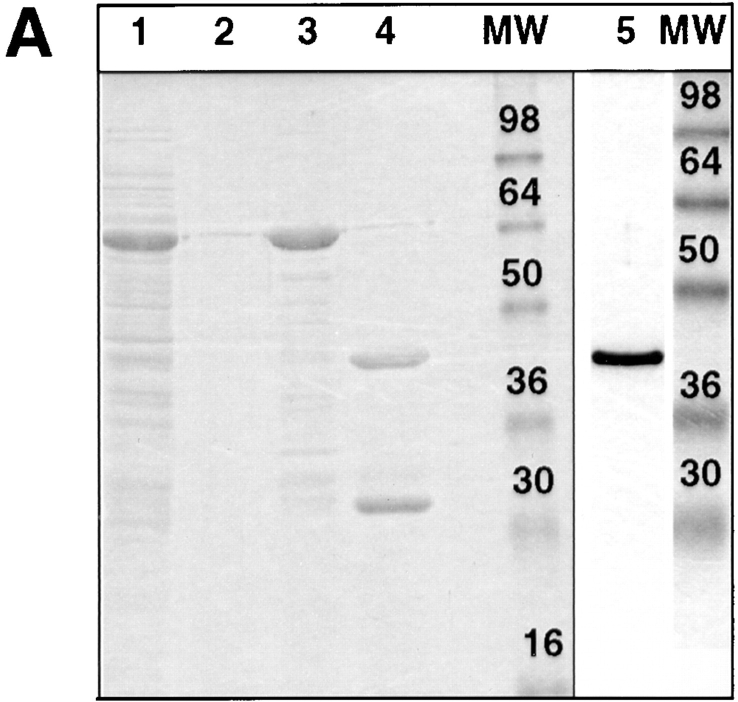
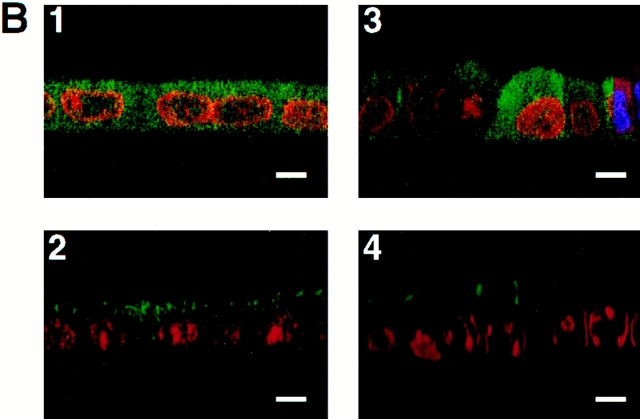
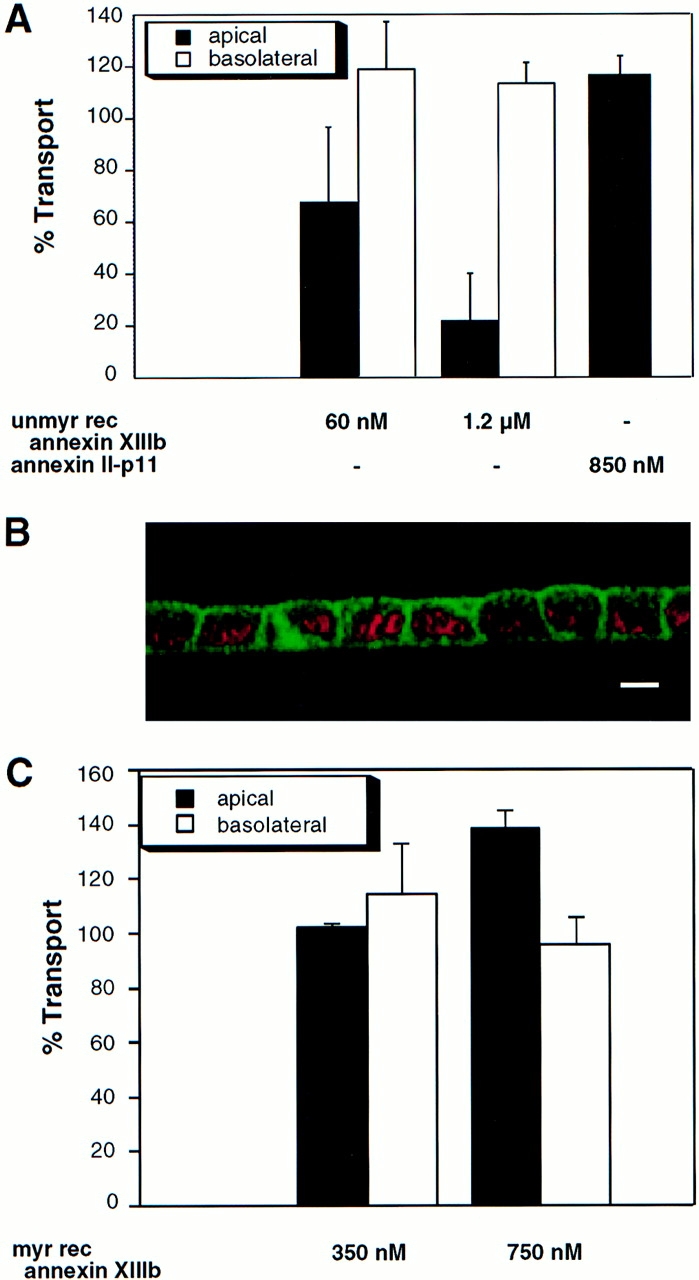
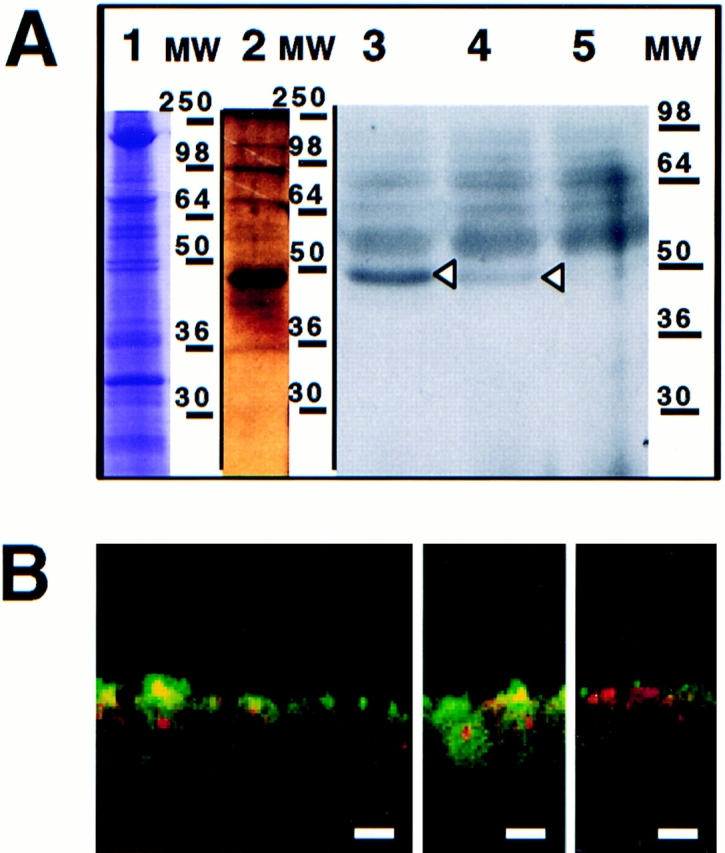
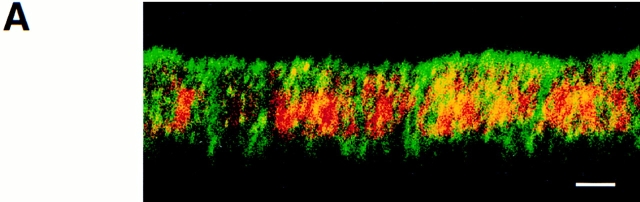
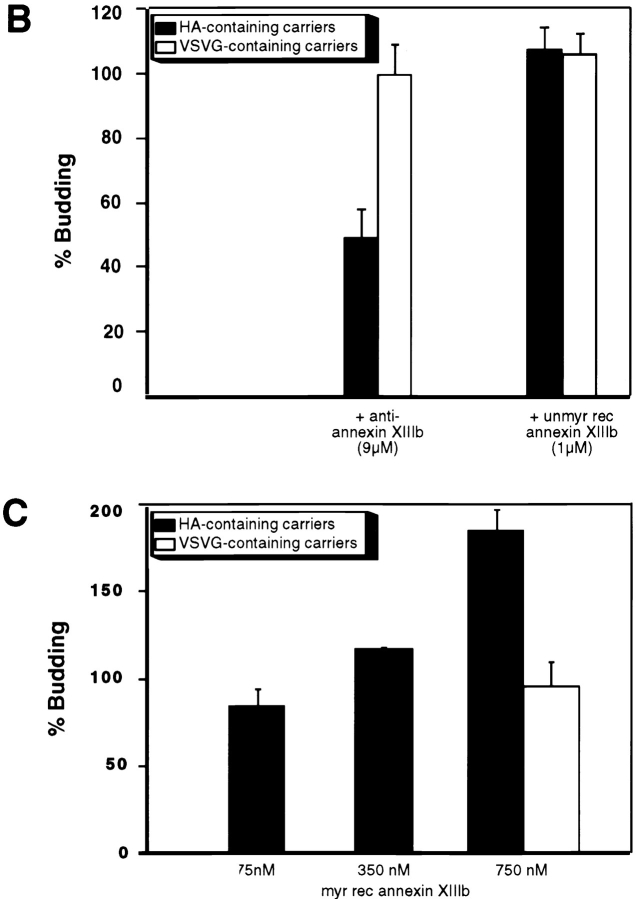
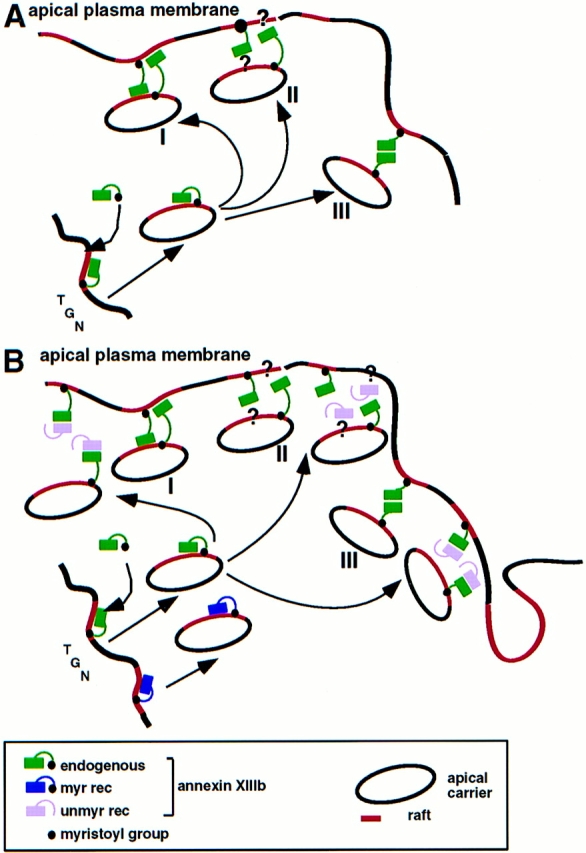
A member of the annexin XIII sub-family, annexin XIIIb, has been implicated in the apical exocytosis of epithelial kidney cells. Annexins are phospholipid-binding proteins that have been suggested to be involved in membrane trafficking events although their actual physiological function remains open. Unlike the other annexins, annexin XIIIs are myristoylated. Here, we show by immunoelectron microscopy that annexin XIIIb is localized to the trans-Golgi network (TGN), vesicular carriers and the apical cell surface. Polarized apical sorting involves clustering of apical proteins into dynamic sphingolipid-cholesterol rafts. We now provide evidence for the raft association of annexin XIIIb. Using in vitro assays and either myristoylated or unmyristoylated recombinant annexin XIIIb, we demonstrate that annexin XIIIb in its native myristoylated form stimulates specifically apical transport whereas the unmyristoylated form inhibits this route. Moreover, we show that formation of apical carriers from the TGN is inhibited by an anti-annexin XIIIb antibody whereas it is stimulated by myristoylated recombinant annexin XIIIb. These results suggest that annexin XIIIb directly participates in apical delivery.
Figures
Similar articles
-
Annexin XIIIb: a novel epithelial specific annexin is implicated in vesicular traffic to the apical plasma membrane.J Cell Biol. 1995 Mar;128(6):1043-53. doi: 10.1083/jcb.128.6.1043. J Cell Biol. 1995. PMID: 7896870 Free PMC article.
-
Annexin XIIIb guides raft-dependent and -independent apical traffic in MDCK cells.Eur J Cell Biol. 2010 Nov;89(11):799-806. doi: 10.1016/j.ejcb.2010.06.022. Epub 2010 Aug 13. Eur J Cell Biol. 2010. PMID: 20708818
-
Different properties of two isoforms of annexin XIII in MDCK cells.J Cell Sci. 2000 Jul;113 ( Pt 14):2607-18. doi: 10.1242/jcs.113.14.2607. J Cell Sci. 2000. PMID: 10862718
-
Mechanisms and functional features of polarized membrane traffic in epithelial and hepatic cells.Biochem J. 1998 Dec 1;336 ( Pt 2)(Pt 2):257-69. doi: 10.1042/bj3360257. Biochem J. 1998. PMID: 9820799 Free PMC article. Review.
-
Polarized sorting in epithelial cells: raft clustering and the biogenesis of the apical membrane.J Cell Sci. 2004 Dec 1;117(Pt 25):5955-64. doi: 10.1242/jcs.01596. J Cell Sci. 2004. PMID: 15564373 Review.
Cited by
-
Annexins as organizers of cholesterol- and sphingomyelin-enriched membrane microdomains in Niemann-Pick type C disease.Cell Mol Life Sci. 2012 Jun;69(11):1773-85. doi: 10.1007/s00018-011-0894-0. Epub 2011 Dec 13. Cell Mol Life Sci. 2012. PMID: 22159585 Free PMC article. Review.
-
Polarized sorting and trafficking in epithelial cells.Cell Res. 2012 May;22(5):793-805. doi: 10.1038/cr.2012.64. Epub 2012 Apr 24. Cell Res. 2012. PMID: 22525333 Free PMC article. Review.
-
Influenza virus assembly and lipid raft microdomains: a role for the cytoplasmic tails of the spike glycoproteins.J Virol. 2000 May;74(10):4634-44. doi: 10.1128/jvi.74.10.4634-4644.2000. J Virol. 2000. PMID: 10775599 Free PMC article.
-
The inducible amphisome isolates viral hemagglutinin and defends against influenza A virus infection.Nat Commun. 2020 Jan 9;11(1):162. doi: 10.1038/s41467-019-13974-w. Nat Commun. 2020. PMID: 31919357 Free PMC article.
-
Cholesterol depletion reduces apical transport capacity in epithelial Madin-Darby canine kidney cells.Biochem J. 2001 Jul 1;357(Pt 1):11-5. doi: 10.1042/0264-6021:3570011. Biochem J. 2001. PMID: 11415430 Free PMC article.
References
-
- Ahmed SN, Brown DA, London E. On the origin of sphingolipid/ cholesterol-rich detergent-insoluble cell membranes: physiological concentrations of cholesterol and sphingolipid induce formation of a detergent-insoluble, liquid-ordered lipid phase in model. Biochemistry. 1997;36:10944–10953. - PubMed
-
- Ali SM, Geisow MJ, Burgoyne RD. A role for calpactin in calcium-dependent exocytosis in adrenal chromaffin cells. Nature. 1989;340:313–315. - PubMed
-
- Beckers CJM, Keller DS, Balch WE. Semi-intact cells permeable to macromolecules: use in reconstitution of protein transport from the endoplasmic reticulum to the golgi complex. Cell. 1987;50:523–534. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
