

Different cis-regulatory DNA elements mediate developmental stage- and tissue-specific expression of the human COL2A1 gene in transgenic mice
- PMID: 9628886
- PMCID: PMC2132792
- DOI: 10.1083/jcb.141.6.1291
Different cis-regulatory DNA elements mediate developmental stage- and tissue-specific expression of the human COL2A1 gene in transgenic mice
Abstract
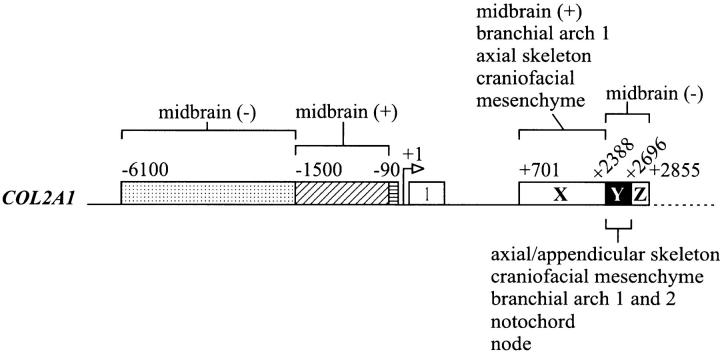
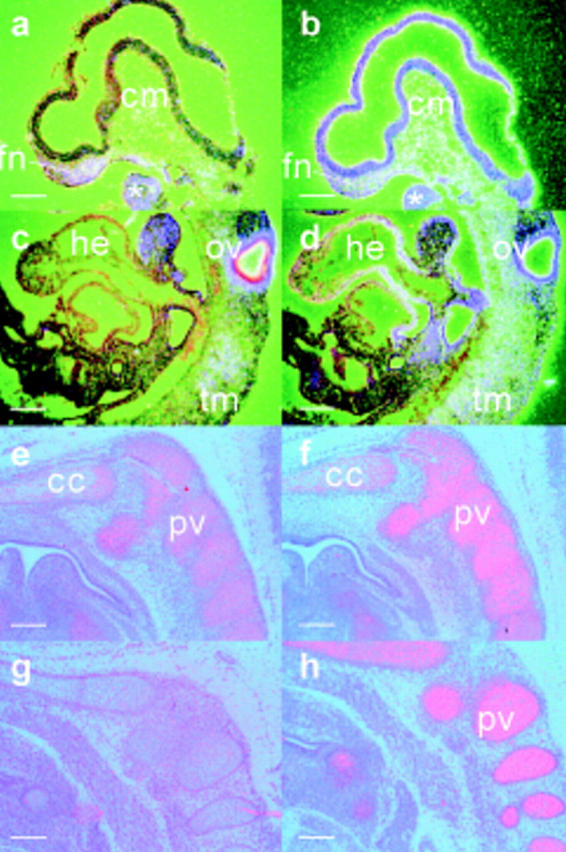
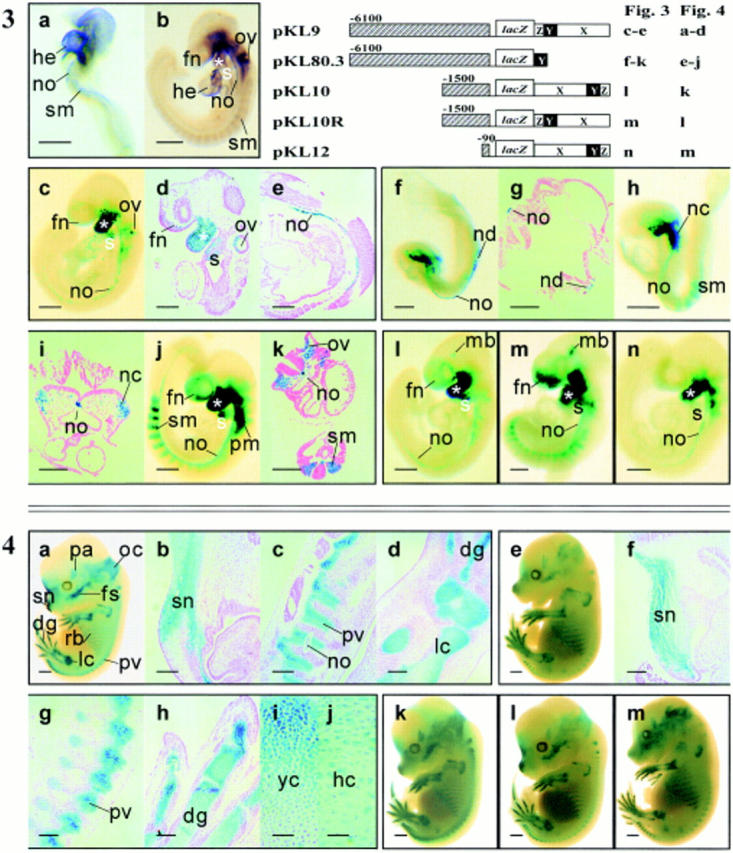
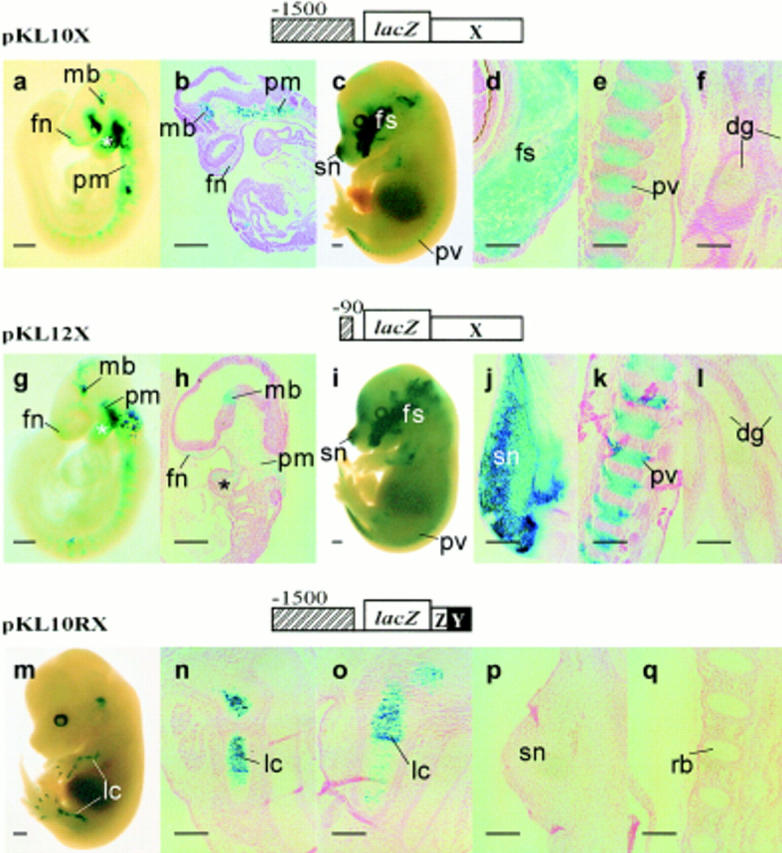
Expression of the type II collagen gene (human COL2A1, mouse Col2a1) heralds the differentiation of chondrocytes. It is also expressed in progenitor cells of some nonchondrogenic tissues during embryogenesis. DNA sequences in the 5' flanking region and intron 1 are known to control tissue-specific expression in vitro, but the regulation of COL2A1 expression in vivo is not clearly understood. We have tested the regulatory activity of DNA sequences from COL2A1 on the expression of a lacZ reporter gene in transgenic mice. We have found that type II collagen characteristic expression of the transgene requires the enhancer activity of a 309-bp fragment (+2, 388 to +2,696) in intron 1 in conjunction with 6.1-kb 5' sequences. Different regulatory elements were found in the 1.6-kb region (+701 to +2,387) of intron 1 which only needs 90-bp 5' sequences for tissue-specific expression in different components of the developing cartilaginous skeleton. Distinct positive and negative regulatory elements act together to control tissue-specific transgene expression in the developing midbrain neuroepithelium. Positive elements affecting expression in the midbrain were found in the region from -90 to -1,500 and from +701 to +2,387, whereas negatively acting elements were detected in the regions from -1,500 to -6,100 and +2,388 to +2,855.
Figures

Similar articles
-
Separable cis-regulatory elements that contribute to tissue- and site-specific alpha 2(XI) collagen gene expression in the embryonic mouse cartilage.J Cell Biol. 1996 Sep;134(6):1573-82. doi: 10.1083/jcb.134.6.1573. J Cell Biol. 1996. PMID: 8830784 Free PMC article.
-
A 182 bp fragment of the mouse pro alpha 1(II) collagen gene is sufficient to direct chondrocyte expression in transgenic mice.J Cell Sci. 1995 Dec;108 ( Pt 12):3677-84. doi: 10.1242/jcs.108.12.3677. J Cell Sci. 1995. PMID: 8719874
-
Regulatory elements of Xenopus col2a1 drive cartilaginous gene expression in transgenic frogs.Int J Dev Biol. 2010;54(1):141-50. doi: 10.1387/ijdb.092848rk. Int J Dev Biol. 2010. PMID: 19757383
-
A new approach for studying gene regulation by distant DNA elements in transgenic mice.Scand J Clin Lab Invest Suppl. 1999;229:33-9. doi: 10.1080/00365519950185931. Scand J Clin Lab Invest Suppl. 1999. PMID: 10097288 Review.
-
Regulation of expression of the alpha 1 (I) collagen gene: a critical appraisal of the role of the first intron.Matrix Biol. 1996 Apr;15(1):3-10. doi: 10.1016/s0945-053x(96)90121-3. Matrix Biol. 1996. PMID: 8783182 Review.
Cited by
-
Joint reconstruction of cis-regulatory interaction networks across multiple tissues using single-cell chromatin accessibility data.Brief Bioinform. 2021 May 20;22(3):bbaa120. doi: 10.1093/bib/bbaa120. Brief Bioinform. 2021. PMID: 32578841 Free PMC article.
-
Brain-specific deletion of extracellular signal-regulated kinase 2 mitogen-activated protein kinase leads to aberrant cortical collagen deposition.Am J Pathol. 2009 Dec;175(6):2586-99. doi: 10.2353/ajpath.2009.090130. Epub 2009 Nov 5. Am J Pathol. 2009. PMID: 19893051 Free PMC article.
-
Discovery and validation of information theory-based transcription factor and cofactor binding site motifs.Nucleic Acids Res. 2017 Mar 17;45(5):e27. doi: 10.1093/nar/gkw1036. Nucleic Acids Res. 2017. PMID: 27899659 Free PMC article.
-
ELF3 modulates type II collagen gene (COL2A1) transcription in chondrocytes by inhibiting SOX9-CBP/p300-driven histone acetyltransferase activity.Connect Tissue Res. 2017 Jan;58(1):15-26. doi: 10.1080/03008207.2016.1200566. Epub 2016 Jun 16. Connect Tissue Res. 2017. PMID: 27310669 Free PMC article.
-
Up-regulation of type II collagen gene by 17β-estradiol in articular chondrocytes involves Sp1/3, Sox-9, and estrogen receptor α.J Mol Med (Berl). 2014 Nov;92(11):1179-200. doi: 10.1007/s00109-014-1195-5. Epub 2014 Aug 2. J Mol Med (Berl). 2014. PMID: 25081415
References
-
- Bell DM, Leung KK, Wheatley SC, Ng LJ, Zhou S, Ling KW, Sham MH, Koopman P, Tam PP, Cheah KS. SOX9 directly regulates the type-II collagen gene. Nat Genet. 1997;16:174–178. - PubMed
-
- Cheah KS, Lau ET, Au PK, Tam PP. Expression of the mouse alpha 1(II) collagen gene is not restricted to cartilage during development. Development (Camb) 1991;111:945–953. - PubMed
-
- Darnell JE. Variety in the level of gene control in eukaryotic cells. Nature. 1982;297:365–371. - PubMed
