

Protein serine/threonine phosphatase Ptc2p negatively regulates the unfolded-protein response by dephosphorylating Ire1p kinase
- PMID: 9528768
- PMCID: PMC121426
- DOI: 10.1128/MCB.18.4.1967
Protein serine/threonine phosphatase Ptc2p negatively regulates the unfolded-protein response by dephosphorylating Ire1p kinase
Abstract
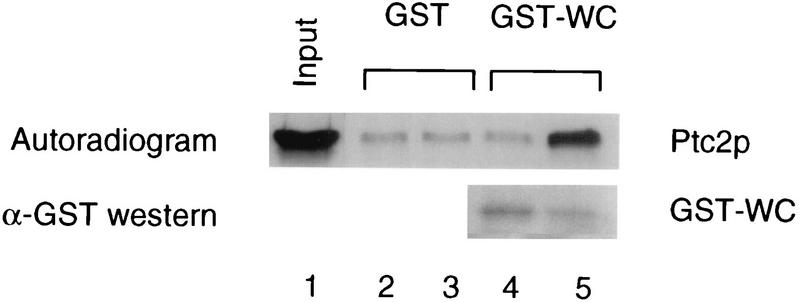
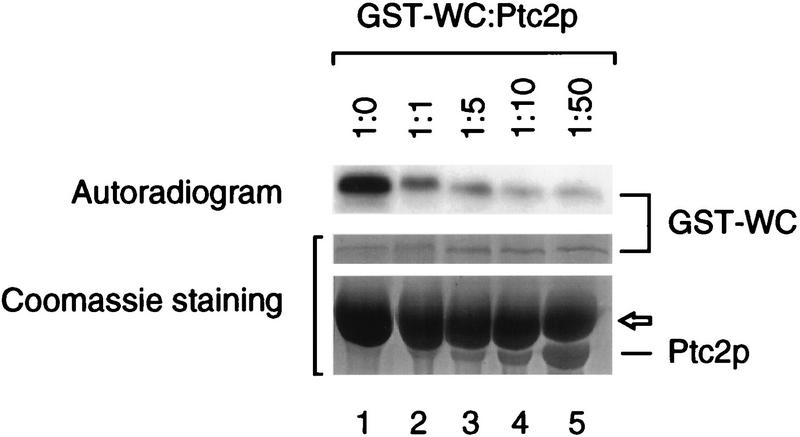
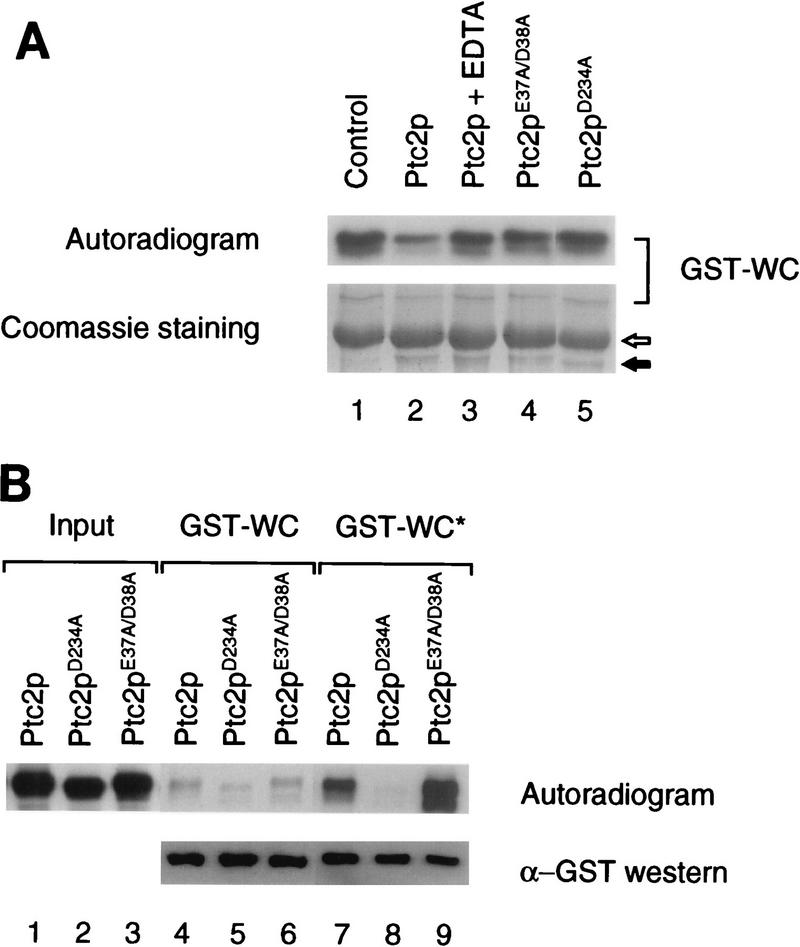
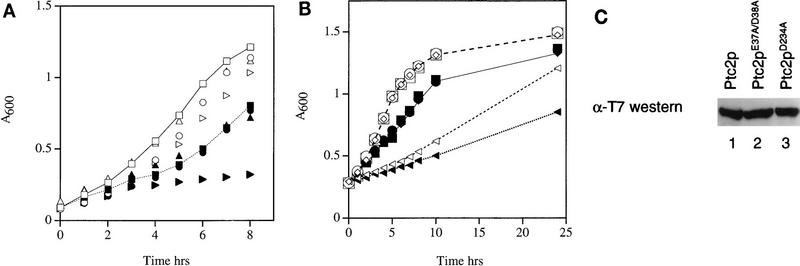
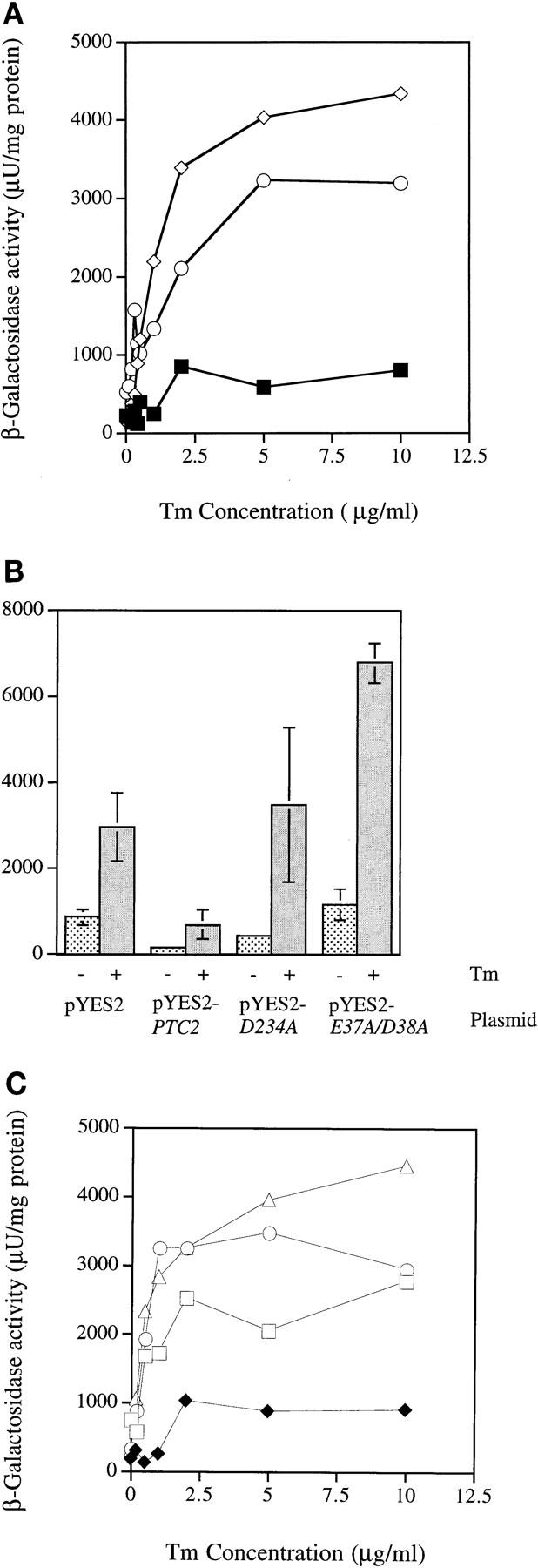
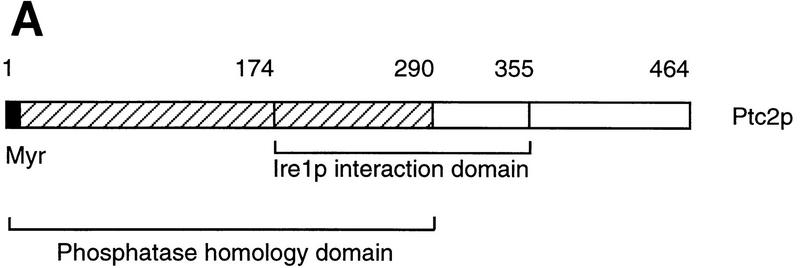
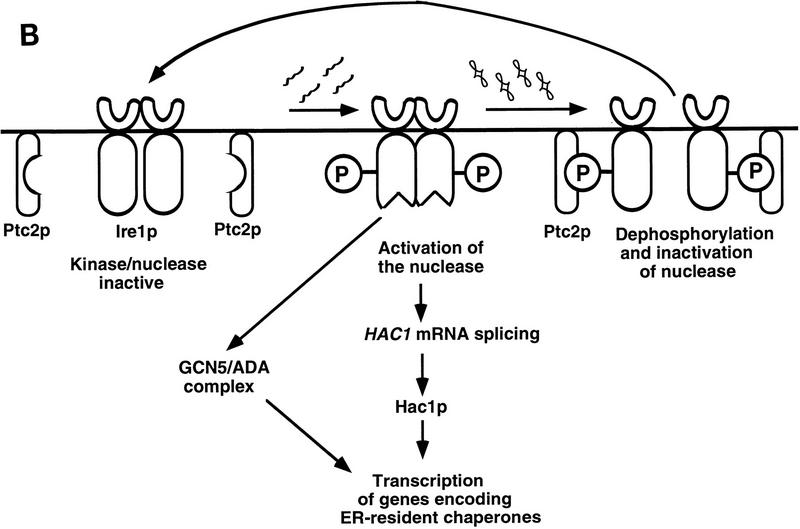
Cells respond to the accumulation of unfolded proteins in the endoplasmic reticulum (ER) by increasing the transcription of the genes encoding ER-resident chaperone proteins. Ire1p is a transmembrane protein kinase that transmits the signal from unfolded proteins in the lumen of the ER by a mechanism that requires oligomerization and trans-autophosphorylation of its cytoplasmic-nucleoplasmic kinase domain. Activation of Ire1p induces a novel spliced form of HAC1 mRNA that produces Hac1p, a transcription factor that is required for activation of the transcription of genes under the control of the unfolded-protein response (UPR) element. Searching for proteins that interact with Ire1p in Saccharomyces cerevisiae, we isolated PTC2, which encodes a serine/threonine phosphatase of type 2C. The Ptc2p interaction with Ire1p is specific, direct, dependent on Ire1p phosphorylation, and mediated through a kinase interaction domain within Ptc2p. Ptc2p dephosphorylates Ire1p efficiently in an Mg2+-dependent manner in vitro. PTC2 is nonessential for growth and negatively regulates the UPR pathway. Strains carrying null alleles of PTC2 have a three- to fourfold-increased UPR and increased levels of spliced HAC1 mRNA. Overexpression of wild-type Ptc2p but not catalytically inactive Ptc2p reduces levels of spliced HAC1 mRNA and attenuates the UPR, demonstrating that the phosphatase activity of Ptc2p is required for regulation of the UPR. These results demonstrate that Ptc2p downregulates the UPR by dephosphorylating Ire1p and reveal a novel mechanism of regulation in the UPR pathway upstream of the HAC1 mRNA splicing event.
Figures



Similar articles
-
The transmembrane kinase Ire1p is a site-specific endonuclease that initiates mRNA splicing in the unfolded protein response.Cell. 1997 Sep 19;90(6):1031-9. doi: 10.1016/s0092-8674(00)80369-4. Cell. 1997. PMID: 9323131
-
A stress response pathway from the endoplasmic reticulum to the nucleus requires a novel bifunctional protein kinase/endoribonuclease (Ire1p) in mammalian cells.Genes Dev. 1998 Jun 15;12(12):1812-24. doi: 10.1101/gad.12.12.1812. Genes Dev. 1998. PMID: 9637683 Free PMC article.
-
The cellular response to protein misfolding in the endoplasmic reticulum.Gene Expr. 1999;7(4-6):293-300. Gene Expr. 1999. PMID: 10440230 Free PMC article. Review.
-
The transcriptional co-activator ADA5 is required for HAC1 mRNA processing in vivo.J Biol Chem. 2000 Feb 4;275(5):3377-81. doi: 10.1074/jbc.275.5.3377. J Biol Chem. 2000. PMID: 10652329
-
Translation Control of HAC1 by Regulation of Splicing in Saccharomyces cerevisiae.Int J Mol Sci. 2019 Jun 12;20(12):2860. doi: 10.3390/ijms20122860. Int J Mol Sci. 2019. PMID: 31212749 Free PMC article. Review.
Cited by
-
Fusel alcohols regulate translation initiation by inhibiting eIF2B to reduce ternary complex in a mechanism that may involve altering the integrity and dynamics of the eIF2B body.Mol Biol Cell. 2010 Jul 1;21(13):2202-16. doi: 10.1091/mbc.e09-11-0962. Epub 2010 May 5. Mol Biol Cell. 2010. PMID: 20444979 Free PMC article.
-
TGFβ1 regulates HRas-mediated activation of IRE1α through the PERK-RPAP2 axis in keratinocytes.Mol Carcinog. 2022 Oct;61(10):958-971. doi: 10.1002/mc.23453. Epub 2022 Aug 17. Mol Carcinog. 2022. PMID: 35975910 Free PMC article.
-
Mammalian transcription factor ATF6 is synthesized as a transmembrane protein and activated by proteolysis in response to endoplasmic reticulum stress.Mol Biol Cell. 1999 Nov;10(11):3787-99. doi: 10.1091/mbc.10.11.3787. Mol Biol Cell. 1999. PMID: 10564271 Free PMC article.
-
The ire1 and ptc2 genes involved in the unfolded protein response pathway in the filamentous fungus Trichoderma reesei.Mol Genet Genomics. 2004 Nov;272(4):443-51. doi: 10.1007/s00438-004-1070-0. Epub 2004 Oct 8. Mol Genet Genomics. 2004. PMID: 15480788
-
Unfolded protein responses with or without unfolded proteins?Cells. 2012 Nov 1;1(4):926-50. doi: 10.3390/cells1040926. Cells. 2012. PMID: 24710536 Free PMC article.
References
-
- Barford D. Molecular mechanisms of the protein serine/threonine phosphatases. Trends Biochem Sci. 1996;21:407–412. - PubMed
-
- Berben G, Dumont J, Gilliquet V, Bolle P A, Hilger F. The YDp plasmids: a uniform set of vectors bearing versatile gene disruption cassettes for Saccharomyces cerevisiae. Yeast. 1991;7:475–477. - PubMed
-
- Boeke J D, LaCroute F, Fink G R. A positive selection for mutants lacking orotidine-5′-phosphate decarboxylase activity in yeast: 5-fluoro-orotic acid resistance. Mol Gen Genet. 1984;197:345–346. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases