

Activation of the mitogen-activated protein kinase/extracellular signal-regulated kinase pathway by conventional, novel, and atypical protein kinase C isotypes
- PMID: 9447975
- PMCID: PMC108790
- DOI: 10.1128/MCB.18.2.790
Activation of the mitogen-activated protein kinase/extracellular signal-regulated kinase pathway by conventional, novel, and atypical protein kinase C isotypes
Abstract
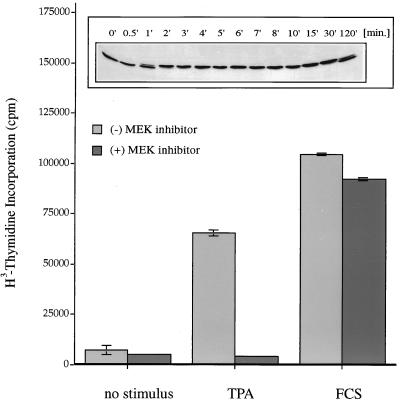
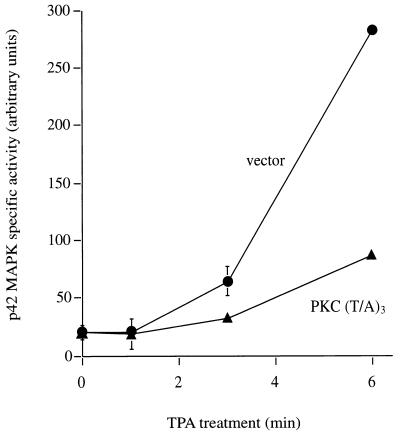
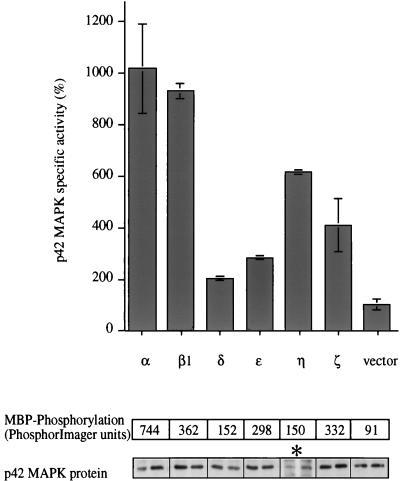
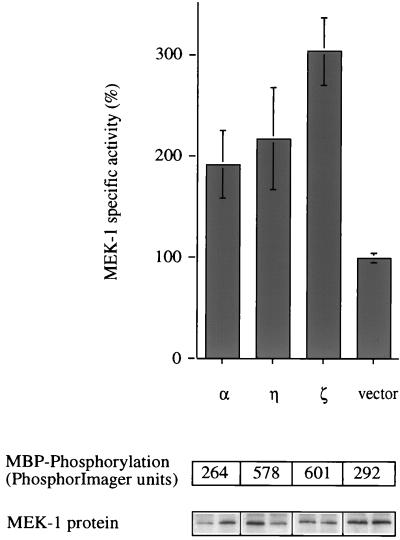
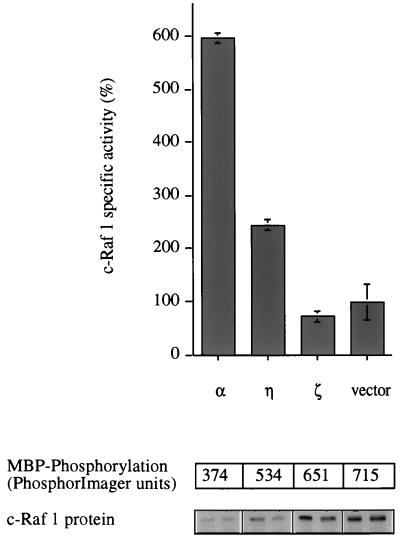
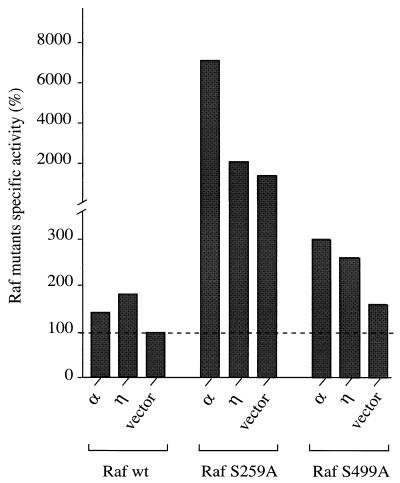
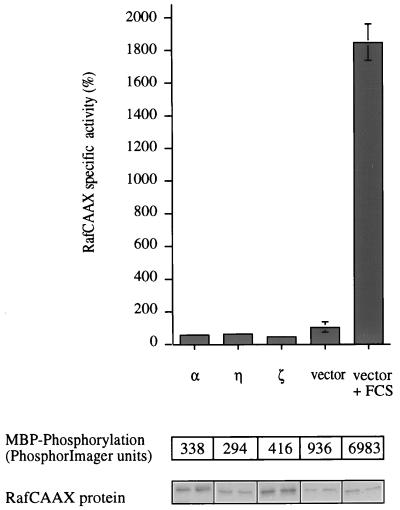
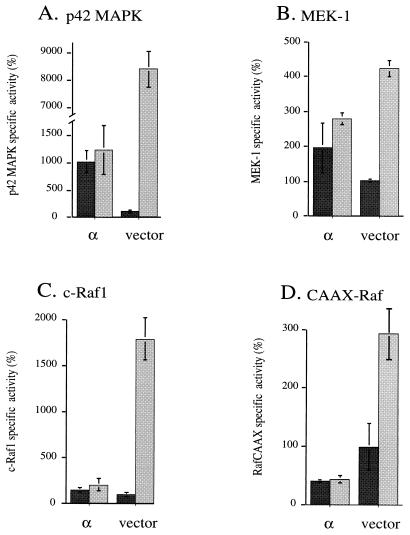
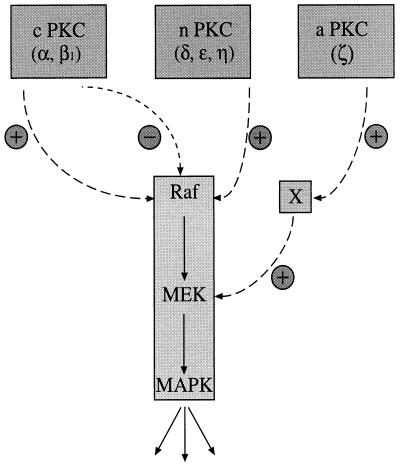
Phorbol ester treatment of quiescent Swiss 3T3 cells leads to cell proliferation, a response thought to be mediated by protein kinase C (PKC), the major cellular receptor for this class of agents. We demonstrate here that this proliferation is dependent on the activation of the extracellular signal-regulated kinase/mitogen-activated protein kinase (ERK/MAPK) cascade. It is shown that dominant-negative PKC-alpha inhibits stimulation of the ERK/MAPK pathway by phorbol esters in Cos-7 cells, demonstrating a role for PKC in this activation. To assess the potential specificity of PKC isotypes mediating this process, constitutively active mutants of six PKC isotypes (alpha, beta, delta, epsilon, eta, and zeta) were employed. Transient transfection of these PKC mutants into Cos-7 cells showed that members of all three groups of PKC (conventional, novel, and atypical) are able to activate p42 MAPK as well as its immediate upstream activator, the MAPK/ERK kinase MEK-1. At the level of Raf, the kinase that phosphorylates MEK-1, the activation cascade diverges; while conventional and novel PKCs (isotypes alpha and eta) are potent activators of c-Raf1, atypical PKC-zeta cannot increase c-Raf1 activity, stimulating MEK by an independent mechanism. Stimulation of c-Raf1 by PKC-alpha and PKC-eta was abrogated for RafCAAX, which is a membrane-localized, partially active form of c-Raf1. We further established that activation of Raf is independent of phosphorylation at serine residues 259 and 499. In addition to activation, we describe a novel Raf desensitization induced by PKC-alpha, which acts to prevent further Raf stimulation by growth factors. The results thus demonstrate a necessary role for PKC and p42 MAPK activation in 12-O-tetradecanoylphorbol-13-acetate induced mitogenesis and provide evidence for multiple PKC controls acting on this MAPK cascade.
Figures
Similar articles
-
Protein kinase C activates the MEK-ERK pathway in a manner independent of Ras and dependent on Raf.J Biol Chem. 1996 Sep 20;271(38):23512-9. doi: 10.1074/jbc.271.38.23512. J Biol Chem. 1996. PMID: 8798560
-
Novel roles of specific isoforms of protein kinase C in activation of the c-fos serum response element.Mol Cell Biol. 1999 Feb;19(2):1313-24. doi: 10.1128/MCB.19.2.1313. Mol Cell Biol. 1999. PMID: 9891065 Free PMC article.
-
Regulation of BCR- and PKC/Ca(2+)-mediated activation of the Raf1/MEK/MAPK pathway by protein-tyrosine kinase and -tyrosine phosphatase activities.Mol Immunol. 1996 Feb;33(3):287-96. doi: 10.1016/0161-5890(95)00134-4. Mol Immunol. 1996. PMID: 8649450
-
Signalling by protein kinase C isoforms in the heart.Mol Cell Biochem. 1996 Apr 12-26;157(1-2):65-72. doi: 10.1007/BF00227882. Mol Cell Biochem. 1996. PMID: 8739230 Review.
-
The MAPK signaling cascade.FASEB J. 1995 Jun;9(9):726-35. FASEB J. 1995. PMID: 7601337 Review.
Cited by
-
Exercise Activates p53 and Negatively Regulates IGF-1 Pathway in Epidermis within a Skin Cancer Model.PLoS One. 2016 Aug 10;11(8):e0160939. doi: 10.1371/journal.pone.0160939. eCollection 2016. PLoS One. 2016. PMID: 27509024 Free PMC article.
-
Protein kinase C isozymes regulate matrix metalloproteinase-1 expression and cell invasion in Helicobacter pylori infection.Gut. 2013 Mar;62(3):358-67. doi: 10.1136/gutjnl-2012-302103. Epub 2012 Mar 22. Gut. 2013. PMID: 22442164 Free PMC article.
-
Atypical PKCiota contributes to poor prognosis through loss of apical-basal polarity and cyclin E overexpression in ovarian cancer.Proc Natl Acad Sci U S A. 2005 Aug 30;102(35):12519-24. doi: 10.1073/pnas.0505641102. Epub 2005 Aug 22. Proc Natl Acad Sci U S A. 2005. PMID: 16116079 Free PMC article.
-
ATP induces synaptic gene expressions in cortical neurons: transduction and transcription control via P2Y1 receptors.Mol Pharmacol. 2010 Dec;78(6):1059-71. doi: 10.1124/mol.110.066506. Epub 2010 Sep 16. Mol Pharmacol. 2010. PMID: 20847060 Free PMC article.
-
C-terminal fragment of tetanus toxin heavy chain activates Akt and MEK/ERK signalling pathways in a Trk receptor-dependent manner in cultured cortical neurons.Biochem J. 2003 Jul 15;373(Pt 2):613-20. doi: 10.1042/BJ20030333. Biochem J. 2003. PMID: 12710887 Free PMC article.
References
-
- Adams P D, Parker P J. TPA-induced activation of MAP kinase. FEBS Lett. 1991;290:77–82. - PubMed
-
- Alessi D R, Cuenda A, Cohen P, Dudley D T, Saltiel A R. PD 098059 is a specific inhibitor of the activation of mitogen-activated protein kinase kinase in vitro and in vivo. J Biol Chem. 1995;270:27489–27494. - PubMed
-
- Avruch J, Zhang X F, Kyriakis J M. Raf meets Ras: completing the framework of a signal transduction pathway. Trends Biochem Sci. 1994;19:279–283. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous