

Expression of foreign proteins by poliovirus polyprotein fusion: analysis of genetic stability reveals rapid deletions and formation of cardioviruslike open reading frames
- PMID: 9420196
- PMCID: PMC109345
- DOI: 10.1128/JVI.72.1.20-31.1998
Expression of foreign proteins by poliovirus polyprotein fusion: analysis of genetic stability reveals rapid deletions and formation of cardioviruslike open reading frames
Abstract
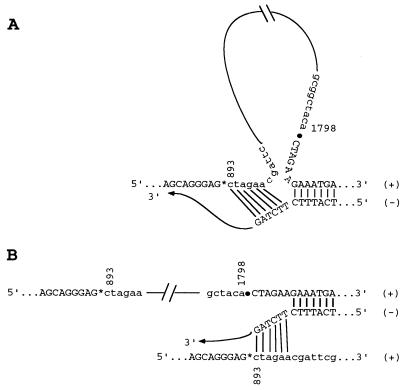
Using a strategy developed by R. Andino, D. Silvera, S. D. Suggett, P. I. Achacoso, C. J. Miller, D. Baltimore, and M. B. Feinberg (Science 265:1448-1451, 1994), we constructed recombinant polioviruses by fusing the open reading frame (ORF) of the green fluorescent protein gene (gfp) of Aequorea victoria or the gag gene (encoding p17-p24) of human immunodeficiency virus type 1 (HIV-1) to the N terminus of the poliovirus polyprotein. All poliovirus expression vectors constructed by us and those obtained from Andino et al. were found to be severely impaired in viral replication and genetically unstable. Upon replication, inserted sequences were rapidly deleted as early as the first growth cycle in HeLa cells. However, the vector viruses did not readily revert to the wild-type sequence but rather retained some of the insert plus the artificial 3Cpro/3CDpro cleavage site, engineered between the heterologous sequence and the poliovirus polyprotein, to give rise to genotypes reminiscent of cardioviruses. These virus variants that carry a small leader polypeptide were now relatively stable, and they grew better than their progenitor strains. Reverse transcription followed by PCR and sequence analysis of the genomic RNAs reproducibly revealed a few preferred genotypes among the isolated deletion variants. The remaining truncated inserts were retained through subsequent passages. In the immediate vicinity of the deletion borders, we observed short direct sequence repeats that we propose are involved in aligning RNA strands for illegitimate (nonhomologous) RNA recombination during minus-strand synthesis. On the basis of our results, which are at variance with published data, the utility of poliovirus vectors to express proteins > 10 kDa in size through fusion with the polyprotein needs to be reevaluated.
Figures







Similar articles
-
Construction and genetic analysis of dicistronic polioviruses containing open reading frames for epitopes of human immunodeficiency virus type 1 gp120.J Virol. 1995 Aug;69(8):4797-806. doi: 10.1128/JVI.69.8.4797-4806.1995. J Virol. 1995. PMID: 7541843 Free PMC article.
-
Toward a poliovirus-based simian immunodeficiency virus vaccine: correlation between genetic stability and immunogenicity.J Virol. 1997 Oct;71(10):7841-50. doi: 10.1128/JVI.71.10.7841-7850.1997. J Virol. 1997. PMID: 9311872 Free PMC article.
-
Attenuated poliovirus strain as a live vector: expression of regions of rotavirus outer capsid protein VP7 by using recombinant Sabin 3 viruses.J Virol. 1994 Jun;68(6):3925-33. doi: 10.1128/JVI.68.6.3925-3933.1994. J Virol. 1994. PMID: 8189529 Free PMC article.
-
Expression of human immunodeficiency virus type 1 (HIV-1) gag, pol, and env proteins from chimeric HIV-1-poliovirus minireplicons.J Virol. 1991 Jun;65(6):2875-83. doi: 10.1128/JVI.65.6.2875-2883.1991. J Virol. 1991. PMID: 1851859 Free PMC article.
-
Molecular biology and cell-free synthesis of poliovirus.Biologicals. 1993 Dec;21(4):349-56. doi: 10.1006/biol.1993.1095. Biologicals. 1993. PMID: 8024750 Review.
Cited by
-
Evaluating a parainfluenza virus 5-based vaccine in a host with pre-existing immunity against parainfluenza virus 5.PLoS One. 2012;7(11):e50144. doi: 10.1371/journal.pone.0050144. Epub 2012 Nov 20. PLoS One. 2012. PMID: 23185558 Free PMC article.
-
A rapid and versatile reverse genetics approach for generating recombinant positive-strand RNA viruses that use IRES-mediated translation.J Virol. 2024 Mar 19;98(3):e0163823. doi: 10.1128/jvi.01638-23. Epub 2024 Feb 14. J Virol. 2024. PMID: 38353536 Free PMC article.
-
Construction of a recombinant rhinovirus accommodating fluorescent marker expression.Influenza Other Respir Viruses. 2018 Nov;12(6):717-727. doi: 10.1111/irv.12602. Epub 2018 Sep 6. Influenza Other Respir Viruses. 2018. PMID: 30120824 Free PMC article.
-
Expression of an antigenic adenovirus epitope in a group B coxsackievirus.J Virol. 2000 May;74(10):4570-8. doi: 10.1128/jvi.74.10.4570-4578.2000. J Virol. 2000. PMID: 10775593 Free PMC article.
-
SNAP-tagged Chikungunya Virus Replicons Improve Visualisation of Non-Structural Protein 3 by Fluorescence Microscopy.Sci Rep. 2017 Jul 18;7(1):5682. doi: 10.1038/s41598-017-05820-0. Sci Rep. 2017. PMID: 28720784 Free PMC article.
References
-
- Altmeyer R, Murdin A D, Harber J J, Wimmer E. Construction and characterization of a poliovirus/rhinovirus antigenic hybrid. Virology. 1991;184:636–644. - PubMed
-
- Andino, R. Personal communication.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
