

doi: 10.1073/pnas.94.24.12792.
KSR stimulates Raf-1 activity in a kinase-independent manner
Affiliations
- PMID: 9371754
- PMCID: PMC24217
- DOI: 10.1073/pnas.94.24.12792
Item in Clipboard
KSR stimulates Raf-1 activity in a kinase-independent manner
Proc Natl Acad Sci U S A.
.
Erratum in
- Proc Natl Acad Sci U S A 1998 mAR 3;95(5):2714-5
Abstract
Kinase suppressor of Ras (KSR) is an evolutionarily conserved component of Ras-dependent signaling pathways. Here, we find that murine KSR (mKSR1) translocates from the cytoplasm to the plasma membrane in the presence of activated Ras. At the membrane, mKSR1 modulates Ras signaling by enhancing Raf-1 activity in a kinase-independent manner. The activation of Raf-1 is mediated by the mKSR1 cysteine-rich CA3 domain and involves a detergent labile cofactor that is not ceramide. These findings reveal another point of regulation for Ras-mediated signal transduction and further define a noncatalytic role for mKSR1 in the multistep process of Raf-1 activation.
Figures
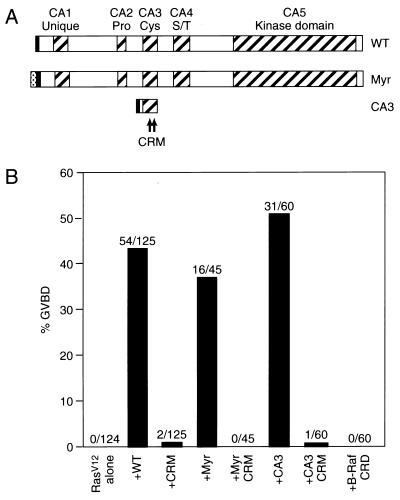
The mKSR1 CA3 domain is required for the augmentation of Ras signaling. (A) Schematic representation of the WT, Myr, and CA3 mKSR1 proteins. The hatched boxes represent the five conserved areas (CA1–CA5) of the KSR family members. Pro, Cys, and S/T indicate that the corresponding CA regions are rich in proline, cysteine, or serine/threonine residues, respectively. The CRM mutation (C359S, C362S) is depicted by asterisks. For Myr mKSR1, the myristylation sequence from the src tyrosine kinase was inserted at the N terminus of WT mKSR1 (gray box). In addition, each construct contained a polyoma-derived (Pyo) epitope tag (solid box). (B) Induction of Xenopus oocyte meiotic maturation by the expression of RasV12 alone or by the coexpression of RasV12 with mKSR1 proteins. GVBD was scored when 0% of the oocytes expressing RasV12 alone and 40% of oocytes coexpressing RasV12 and WT mKSR1 had undergone GVBD. The percentage of oocytes undergoing GVBD is expressed as a solid bar, and the ratio of the number of oocytes undergoing GVBD to the total number injected is displayed above each bar. The numbers obtained represent a compilation of at least three independent experiments where equivalent amounts of the mKSR1 and RasV12 proteins were expressed.

The mKSR1 CA3 domain is required for the Ras-dependent membrane localization of mKSR1. (A) 293 cells transiently expressing WT and Myr mKSR1 or coexpressing WT and RasV12 were fractionated into membrane and cytosolic fractions. The mKSR1 proteins were immunoprecipitated and examined by immunoblot analysis using αPyo antibody. (B) 293 cells coexpressing RasV12 and either WT or CRM mKSR1 proteins were analyzed as in A.

The mKSR1 CA3 domain augments Raf-1 activity in a detergent-sensitive manner. Xenopus oocytes expressing Raf-1 alone (−) or coexpressing Raf-1 and the mKSR1 CA3 domain (+) were injected with RasV12 RNA. Immediately after (0 min) or 150 min after RasV12 injection, oocytes were lysed in hypotonic buffer and membranes were isolated. (A) Raf-1 proteins were immunoprecipitated from membrane fractions resuspended in RIPA buffer (+ detergent) or phosphate-buffered saline (− detergent), and in vitro kinase assays were performed using kinase-inactive MEK as a substrate (1). Phosphorylation of MEK1 on Ser-218 and Ser-222 was determined by tryptic peptide mapping analysis. (B) RasV12 and CA3 proteins were immunoprecipitated from membrane (P100) and cytosolic fractions (S100) isolated at 150 min after injection with RasV12 RNA and were examined by immunoblot analysis using Ras and Pyo antibody, respectively. The migration of processed (Pro) and unprocessed (UnPro) Ras proteins is indicated.
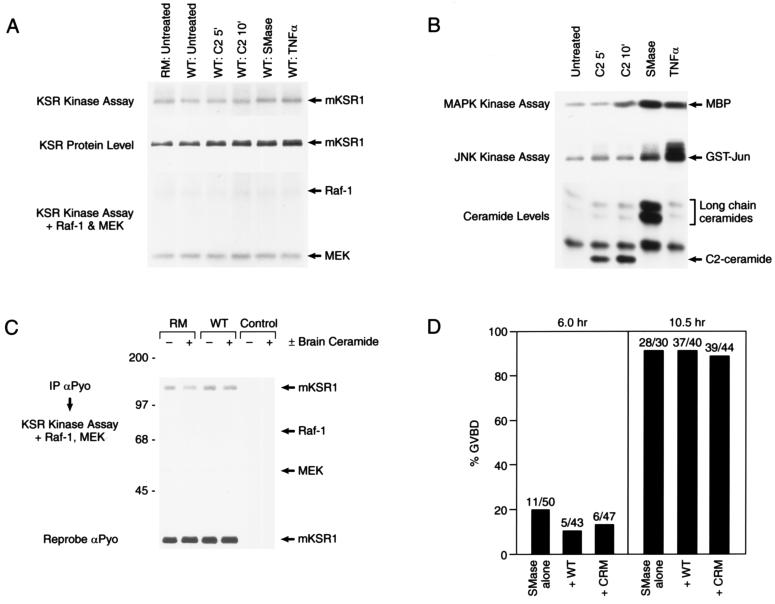
Augmentation of Ras signaling by mKSR1 does not involve ceramide. (A) Cos cells were transiently transfected with constructs encoding wild-type (WT) or kinase-inactive (RM) mKSR1. At 60 hr posttransfection, serum starved cells were left untreated or were stimulated with 20 μM C2 ceramide for 5 or 10 min, 100 milliunits/ml sphingomyelinase (SMase) for 20 min or 10 nM tumor necrosis factor α (TNFα) for 20 min. KSR proteins were immunoprecipitated using αPyo antibody, and mKSR1 immune complex kinase assays were performed in vitro as described by Zhang et al. (ref. ; Top). Immunoprecipitated mKSR1 was detected by immunoblot analysis (Middle). To observe phosphorylation of Raf-1 or modulation of Raf-1 activity, purified activated Raf-1, coexpressed in Sf9 cells in the presence of RasV12 and v-src, and kinase-inactive MEK1 were added to the mKSR1 immune complex kinase assays previously described (ref. ; Bottom). (B) Cos cells were treated as in A and endogenous ceramide levels, JNK activity, and MAPK activity were determined. Ceramide levels were normalized to the untreated control. C2-ceramide levels were elevated 2.9- and 3.7-fold at 5 and 10 min, respectively, and long-chain ceramide levels were elevated 12-fold by SMase and 1.9-fold by TNFα. (C) Purified brain ceramide (100 nM) (+) or diluent (−) was added in vitro to mKSR proteins immunoprecipitated from transfected Cos cells and immune complex kinase assays performed in the presence of activated Raf-1 and kinase-inactive MEK1 as previously described (ref. ; Top). Immunoprecipitated mKSR1 was detected by immunoblot analysis (Middle). (D) Oocytes preinjected with buffer or RNA encoding WT and CRM mKSR1 constructs were treated with 250 milliunits sphingomyelinase. GVBD was then scored 6 and 10.5 hr after treatment.
Similar articles
-
KSR modulates signal propagation within the MAPK cascade.Genes Dev. 1996 Nov 1;10(21):2684-95. doi: 10.1101/gad.10.21.2684. Genes Dev. 1996. PMID: 8946910
-
Solution structure and functional analysis of the cysteine-rich C1 domain of kinase suppressor of Ras (KSR).J Mol Biol. 2002 Jan 18;315(3):435-46. doi: 10.1006/jmbi.2001.5263. J Mol Biol. 2002. PMID: 11786023
-
Kinase suppressor of ras is necessary for tumor necrosis factor alpha activation of extracellular signal-regulated kinase/mitogen-activated protein kinase in intestinal epithelial cells.Cancer Res. 2001 Feb 1;61(3):963-9. Cancer Res. 2001. PMID: 11221891
-
Untying the regulation of the Raf-1 kinase.Arch Biochem Biophys. 2002 Aug 1;404(1):3-9. doi: 10.1016/s0003-9861(02)00244-8. Arch Biochem Biophys. 2002. PMID: 12127063 Review.
-
Ras signaling: PP2A puts Ksr and Raf in the right place.Curr Biol. 2003 Aug 19;13(16):R635-7. doi: 10.1016/s0960-9822(03)00568-2. Curr Biol. 2003. PMID: 12932339 Review. No abstract available.
Cited by
-
Tumor biomarkers for diagnosis, prognosis and targeted therapy.Signal Transduct Target Ther. 2024 May 20;9(1):132. doi: 10.1038/s41392-024-01823-2. Signal Transduct Target Ther. 2024. PMID: 38763973 Free PMC article. Review.
-
Targeting CRAF kinase in anti-cancer therapy: progress and opportunities.Mol Cancer. 2023 Dec 18;22(1):208. doi: 10.1186/s12943-023-01903-x. Mol Cancer. 2023. PMID: 38111008 Free PMC article. Review.
-
Navigating the ERK1/2 MAPK Cascade.Biomolecules. 2023 Oct 20;13(10):1555. doi: 10.3390/biom13101555. Biomolecules. 2023. PMID: 37892237 Free PMC article. Review.
-
Ras Mitogen-activated Protein Kinase Signaling and Kinase Suppressor of Ras as Therapeutic Targets for Hepatocellular Carcinoma.J Liver Cancer. 2021 Mar;21(1):1-11. doi: 10.17998/jlc.21.1.1. Epub 2021 Mar 31. J Liver Cancer. 2021. PMID: 37384270 Free PMC article. Review.
-
Pseudokinase NRP1 facilitates endocytosis of transferrin in the African trypanosome.Sci Rep. 2022 Nov 3;12(1):18572. doi: 10.1038/s41598-022-22054-x. Sci Rep. 2022. PMID: 36329148 Free PMC article.
References
-
- Therrien M, Michaud N R, Rubin G M, Morrison D K. Genes Dev. 1996;10:2684–2695. - PubMed
-
- Therrien M, Chang H C, Solomon N M, Karim F D, Wassarman D A, Rubin G M. Cell. 1995;83:879–888. - PubMed
-
- Sundaram M, Han M. Cell. 1995;83:889–901. - PubMed
-
- Kornfeld K, Hom D B, Horvitz H R. Cell. 1995;83:903–913. - PubMed
-
- Xing H, Kornfeld K, Muslin A J. Curr Biol. 1997;7:294–300. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
