

doi: 10.1073/pnas.94.14.7221.
Slow dimer dissociation of the TATA binding protein dictates the kinetics of DNA binding
Affiliations
- PMID: 9207072
- PMCID: PMC23798
- DOI: 10.1073/pnas.94.14.7221
Item in Clipboard
Slow dimer dissociation of the TATA binding protein dictates the kinetics of DNA binding
Proc Natl Acad Sci U S A.
.
Abstract
The association of the TATA binding protein (TBP) to eukaryotic promoters is a possible rate-limiting step in gene expression. Slow promoter binding might be related to TBP's ability to occlude its DNA binding domain through dimerization. Using a "pull-down" based assay, we find that TBP dimers dissociate slowly (t1/2 = 6-10 min), and thus present a formidable kinetic barrier to TATA binding. At 10 nM, TBP appears to exist as a mixed population of monomers and dimers. In this state, TATA binding displays burst kinetics that appears to reflect rapid binding of monomers and slow dissociation of dimers. The kinetics of the slow phase is in excellent agreement with direct measurements of the kinetics of dimer dissociation.
Figures
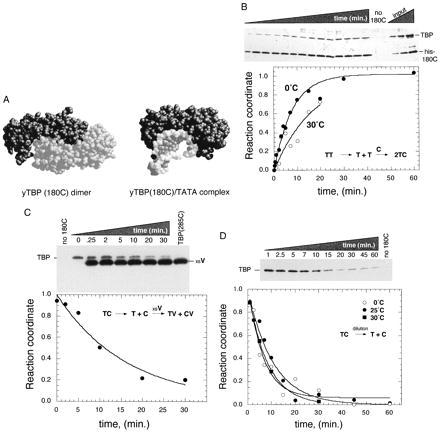
Kinetics of TBP dimer dissociation. (A) Space-filling representations, based upon crystal structure determinations, of the conserved core DNA binding domain of yeast TBP in the absence (Left) and presence (Right) of DNA (13, 14). (B) TBP subunit exchange as measured by the NAB assay (9). •, 1 μM TBP (T) and 2.5 μM his-180C (C) were mixed at 0°C. ○, 1 μM TBP and 0.25 μM his-180C were mixed at 30°C. At various times, samples were quenched and subjected to the NAB assay. A silver-stained gel of the raw 0°C data is shown above the graph of the quantitated data. His-180C was omitted from reactions indicated by “no 180C” (shown in duplicate). Increasing amounts of input proteins, which provide a standard for linearity of the signal, are shown as “input.” (C) TBP subunit exchange measured by dissociation of his-180C/TBP heterodimers. A total of 0.01 μM TBP and 0.25 μM his-180C was mixed and allowed to reach equilibrium at 0°C. A total of 10 μM TBP(285C), designated as “xsV,” was then added to initiate net dissociation of TBP from his-180C. At various times, samples were processed through the NAB assay. A TBP Western blot of the raw data is shown above the quantitation. His-180C was omitted from lanes designated “no 180C.” TBP(285C) was omitted from the 0 min. point. Because TBP(285C) was added at a 1,000-fold excess over TBP, a small but significant amount contaminates the washed resin. The his-180C signal is not shown. (D) Dissociation of his-180C/TBP heterodimers by dilution. TBP (0.025 μM) and his-180C (0.125 μM) were bound to nickel resin at 0°C, allowed to reach equilibrium, then diluted 300-fold into equivalent buffer at 0°C (○), 25°C (•), or 30°C (▪) to initiate net dissociation of TBP dimers. At various times, samples were processed through the NAB assay. A TBP Western blot of the raw data is shown above the quantitation for the 25°C data. Due to the low immunoreactivity of his-180C, its signal was detected at low levels only after extended exposure to film. In all cases, the recovery of his-180C on the resin was constant, indicating stable association of his-180C with the nickel resin (data not shown). In B–D, all TBP band intensities were within the linear range of quantitation.
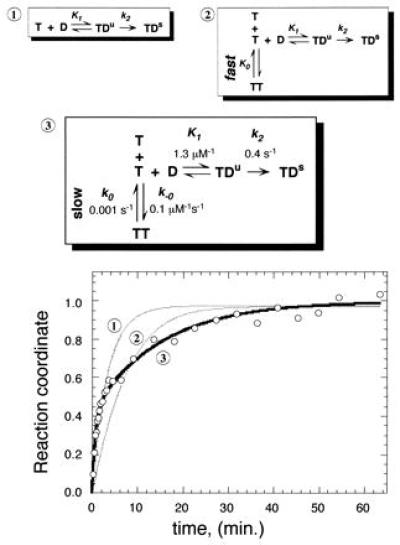
TBP/TATA association displays burst kinetics. Three possible mechanisms for the binding of TBP (T) to TATA (D) are shown. Mechanism 1 includes two steps as proposed by others (20, 21), in which an unstable TBP/TATA complex (TDu) isomerizes into a stable TDs complex. Mechanisms 2 and 3 include TBP dimerization (TT) as a competing reaction, in which dimers are either in rapid equilibrium with monomers (mechanism 2) or represent a kinetically significant step (mechanism 3). TBP was preincubated at 0°C for >60 min. at 10 nM. [32P]TATA DNA (28 bp and 50 nM) was added to initiate the binding time course. This TATA concentration is ≈100-fold higher than the KD for the TBP/TATA complex (23), and is therefore saturating. Data were normalized such that the reaction coordinate ranged from 0 to 1 as described in Materials and Methods. The data were fit to mechanisms 1, 2, and 3 as indicated on the graph using kinsim software (26). Rate constants for a fit to mechanism 3 are shown in the diagram of mechanism 3. The program did not provide error evaluations. Varying any of the kinetic rate constants by more than 10% yielded curves that provided a noticeably poorer fit of the data.
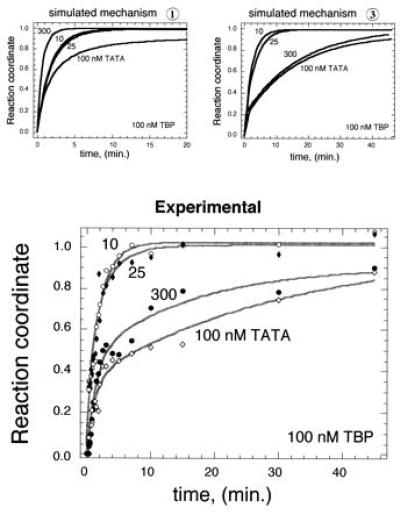
Experimental distinction between mechanisms 1 and 3. Using the rate constants determined in Fig. 2, kinsim was used to simulate binding according to either mechanism 1 or 3 at 100 nM TBP and 10, 25, 100, and 300 nM TATA DNA. Binding reactions were then performed and assayed as described in Fig. 2.
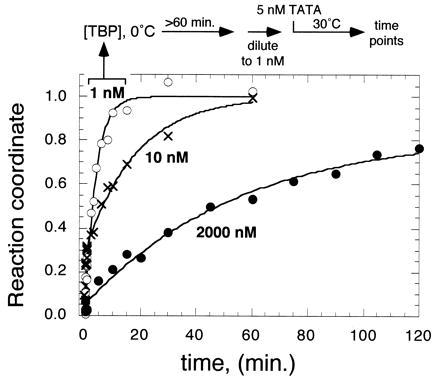
Concentration and preincubation effect on TBP/TATA binding. TBP (1, 10, or 2,000 nM, as indicated) was preincubated for at least 60 min at 0°C. Reactions were then diluted to 1 nM at 30°C, and binding was immediately initiated with the addition of 5 nM [32P]TATA DNA. Binding was assayed as described in Fig. 2.
Similar articles
-
Dimer dissociation and thermosensitivity kinetics of the Saccharomyces cerevisiae and human TATA binding proteins.Biochemistry. 1999 Aug 31;38(35):11340-8. doi: 10.1021/bi990911p. Biochemistry. 1999. PMID: 10471284
-
TFIIA regulates TBP and TFIID dimers.Mol Cell. 1999 Sep;4(3):451-7. doi: 10.1016/s1097-2765(00)80453-0. Mol Cell. 1999. PMID: 10518227
-
Dimerization of the TATA binding protein.J Biol Chem. 1995 Jun 9;270(23):13842-9. doi: 10.1074/jbc.270.23.13842. J Biol Chem. 1995. PMID: 7775442
-
Dynamic interplay of TFIIA, TBP and TATA DNA.J Mol Biol. 1997 Aug 8;271(1):61-75. doi: 10.1006/jmbi.1997.1152. J Mol Biol. 1997. PMID: 9300055
-
X-ray crystallographic studies of eukaryotic transcription initiation factors.Philos Trans R Soc Lond B Biol Sci. 1996 Apr 29;351(1339):483-9. doi: 10.1098/rstb.1996.0046. Philos Trans R Soc Lond B Biol Sci. 1996. PMID: 8735270 Review.
Cited by
-
Structural and functional analysis of mutations along the crystallographic dimer interface of the yeast TATA binding protein.Mol Cell Biol. 2003 May;23(9):3186-201. doi: 10.1128/MCB.23.9.3186-3201.2003. Mol Cell Biol. 2003. PMID: 12697819 Free PMC article.
-
A kinetic model of TBP auto-regulation exhibits bistability.Biol Direct. 2010 Aug 5;5:50. doi: 10.1186/1745-6150-5-50. Biol Direct. 2010. PMID: 20687914 Free PMC article.
-
A single point mutation in TFIIA suppresses NC2 requirement in vivo.EMBO J. 2000 Feb 15;19(4):672-82. doi: 10.1093/emboj/19.4.672. EMBO J. 2000. PMID: 10675336 Free PMC article.
-
Transcription factors that influence RNA polymerases I and II: To what extent is mechanism of action conserved?Biochim Biophys Acta Gene Regul Mech. 2017 Feb;1860(2):246-255. doi: 10.1016/j.bbagrm.2016.10.010. Epub 2016 Oct 27. Biochim Biophys Acta Gene Regul Mech. 2017. PMID: 27989933 Free PMC article. Review.
-
Analysis of homodimer formation in 12-oxophytodienoate reductase 3 in solutio and crystallo challenges the physiological role of the dimer.Sci Rep. 2024 Aug 5;14(1):18093. doi: 10.1038/s41598-024-69160-6. Sci Rep. 2024. PMID: 39103552 Free PMC article.
References
-
- Hernandez N. Genes Dev. 1993;7:1291–1308. - PubMed
-
- Zawel L, Reinberg D. Annu Rev Biochem. 1995;64:533–561. - PubMed
-
- Pugh B F. Curr Opin Cell Biol. 1996;8:303–311. - PubMed
-
- Wang W, Gralla J D, Carey M. Genes Dev. 1992;6:1716–1727. - PubMed
-
- Lieberman P M, Berk A J. Genes Dev. 1994;8:995–1006. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
