

doi: 10.1083/jcb.137.7.1495.
Sec2p mediates nucleotide exchange on Sec4p and is involved in polarized delivery of post-Golgi vesicles
Affiliations
- PMID: 9199166
- PMCID: PMC2137815
- DOI: 10.1083/jcb.137.7.1495
Item in Clipboard
Sec2p mediates nucleotide exchange on Sec4p and is involved in polarized delivery of post-Golgi vesicles
J Cell Biol.
.
Abstract
The small GTPase Sec4p is required for vesicular transport at the post-Golgi stage of yeast secretion. Here we present evidence that mutations in SEC2, itself an essential gene that acts at the same stage of the secretory pathway, cause Sec4p to mislocalize as a result of a random rather than a polarized accumulation of vesicles. Sec2p and Sec4p interact directly, with the nucleotide-free conformation of Sec4p being the preferred state for interaction with Sec2p. Sec2p functions as an exchange protein, catalyzing the dissociation of GDP from Sec4 and promoting the binding of GTP. We propose that Sec2p functions to couple the activation of Sec4p to the polarized delivery of vesicles to the site of exocytosis.
Figures
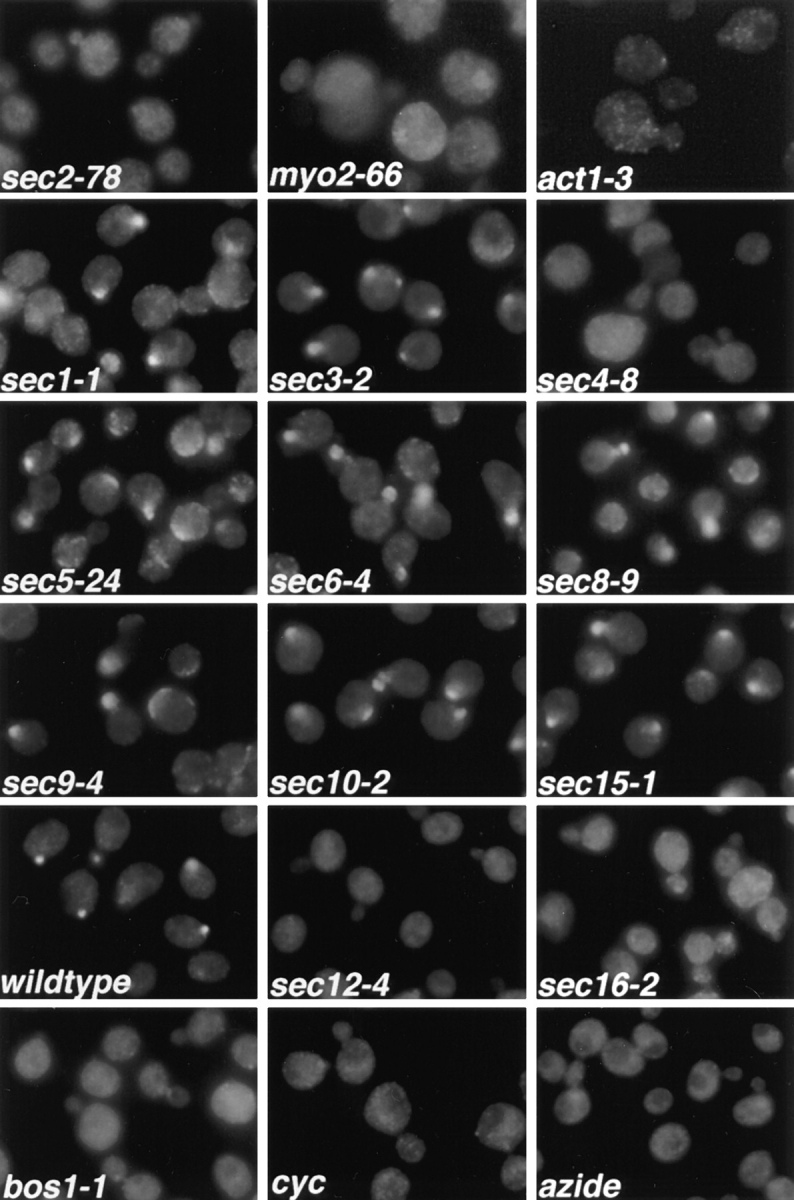
Immunofluorescence localization of
Sec4p in sec mutants. Cells were grown overnight
in YP dextrose medium at 25°C and then shifted
to 37°C for 1 h, fixed, and labeled with αSec4
antibody. The effect of cycloheximide (cyc) and
azide treatment on wild-type cells was tested by
adding either agent to the culture immediately before shift to 37°C. sec2-78, NY1529; myo2-66,
NY1005; act1-3, NY278; sec1-1, NY8; sec3-2,
NY45; sec4-8, NY28; sec5-24, NY30; sec6-4,
NY18; sec8-9, NY43; sec9-4, NY32; sec10-2,
NY36; sec15-1, NY39; wild-type, NY10; sec12-4,
NY738; sec16-2, NY416; bos1-1, NY1282 (see
Table I for strain genotypes). Wild-type cells and
most late-acting sec mutants show immunolabeling preferentially in the bud. In sec2-78, myo2-66, and act1-3, diffuse staining for Sec4p is observed. Also, in sec4-8, ER to Golgi sec mutants
(sec12-4, sec16-2, bos1-1), and after treatment of
wild-type cells with cycloheximide or azide, no
concentration of immunolabeling in the bud is
found.
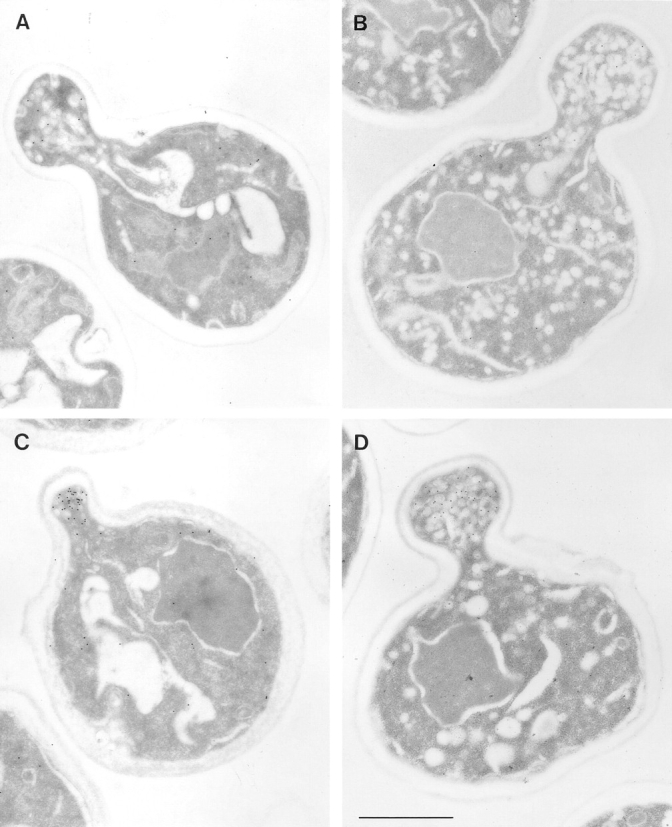
Electron microscopic analysis of sec2-78, wild-type, and sec6-4 cells labeled for Sec4p with immunogold. Cells were grown
overnight at 25°C, shifted to 37°C for 30 min, and then prepared for postembedding immunogold labeling as described in Materials and
Methods. (A) A sec2-78 (NY1529) cell grown at the permissive temperature where some immunogold labeling is found on vesicles in the
bud. Both wild-type (NY10, C) and sec6-4 (NY1294, D) cells show a concentration of immunogold labeling on vesicles in the bud after
shift to the restrictive temperature. In contrast, an average sec2-78 cell in B has immunogold-labeled vesicles randomly distributed in the
bud and mother cell. At 25°C (A) 3 ± 2 gold particles were observed on vesicles while 3 ± 2 gold particles were found independent of
vesicles in the bud. In the mother cell were 1 ± 1 gold particle on vesicles and 10 ± 3 gold particles not on vesicles. At 37°C (B) we found
3 ± 2 gold particles on vesicles and 3 ± 4 gold particles not on vesicles in the bud. In the mother cell were 4 ± 3 gold particles on vesicles
and 15 ± 9 gold particles not on vesicles. The error represents the SD for n = 12 at 25°C and n = 24 at 37°C. Bar, 1 μm.
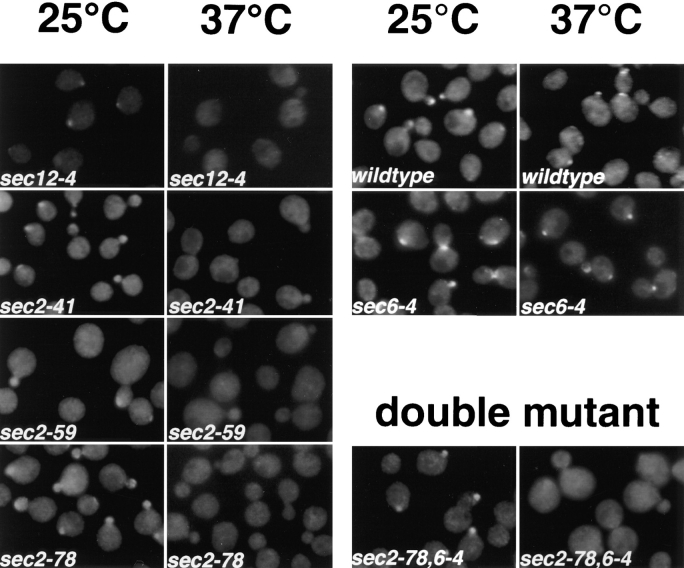
Immunofluorescence localization of Sec4p in
different sec2 alleles and a
sec2-78,6-4 double mutant after a short shift to the restrictive temperature. Cells were
grown overnight in YP dextrose medium at 25°C and
then shifted to 37°C for 10
min, fixed, and labeled with
αSec4 antibody. sec12-4,
NY738; sec2-41, NY132;
sec2-59, NY27; sec2-78,
NY1529; wild-type, NY10;
sec6-4, NY779; sec2-78,6-4,
NY1530. In sec2-41, sec2-59,
and sec2-78, Sec4p, immunostaining was concentrated in
the bud at 25°C but was diffuse already after 10-min
shift to the restrictive temperature. In the ER-to-Golgi
mutant sec12-4, diffuse immunolabeling was also already observed after 10 min
at 37°C. Both in wild-type
and sec6-4, Sec4p immunolabeling was concentrated at
the bud both at 25°C and after 10 min at 37°C. The double mutant was constructed
as described in Materials and
Methods. sec2-78, sec2-78,6-4
shows labeling in the buds at
the permissive temperature
that is no longer detected after 10 min at the restrictive
temperature.
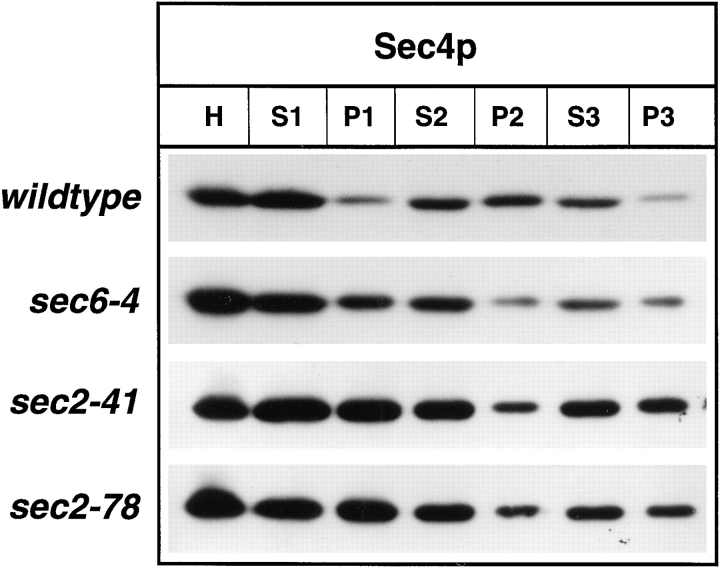
Differential centrifugation of lysates derived from wild-type (NY10), sec6-4 (NY779), sec2-41 (NY132), and sec2-78
(NY1529). Cells were grown overnight in YP dextrose and then
shifted to 37°C. After 2 h at the restrictive temperature, cells
were harvested, and lysates (S1) were prepared as described in
Materials and Methods. S1 supernatants were centrifuged at
10,000 g to generate supernatant S2 and pellet P2. The S2 supernatants were centrifuged at 100,000 g to obtain supernatant S3
and pellet P3. Equal volumes of samples were prepared for electrophoresis, separated on SDS-PAGE gels, and transferred to nitrocellulose. Western blots were probed with αSec4p polyclonal
antibodies and iodinated protein A. No difference in the relative
distribution of Sec4p between S3 and P3 in sec2-41 and sec2-78
compared to sec6-4 is seen. The shift of Sec4p from P2 to P3 in
sec6-4 compared to wild-type is caused by vesicle accumulation.
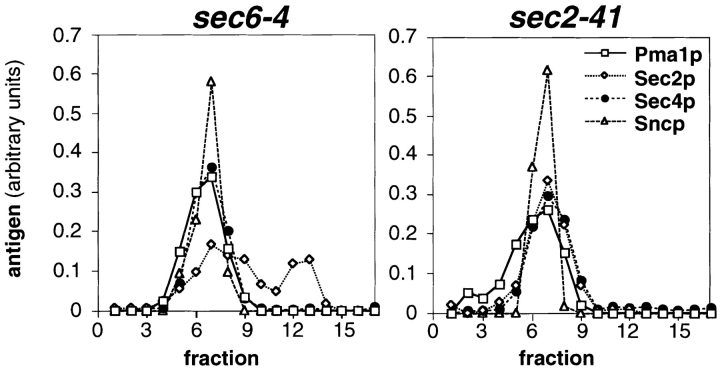
Fractionation of P3 pellets from sec6-4 and sec2-41 cells
shifted to 37°C for 2 h on sucrose density gradients. The P3 pellets were generated by differential centrifugation and loaded in
0.32 M sorbitol on top of 0.4–1.75 M sucrose gradients. After centrifugation to equilibrium, the gradients were fractionated from
the bottom (fraction 2) in 17 fractions (0.75 ml), and the residual
pellets were resuspended in 0.75 ml (fraction 1). Equal volumes
of each fraction were analyzed by SDS-PAGE and immunoblotting (affinity-purified polyclonal antibodies to Pma1p and Sec2p
detected by chemoluminescence; antisera to Sec4p and Sncp detected by iodinated protein A). The Western blots were quantified by densitometry and for each antigen and gradient normalized to a total integrated optical density of 1. In sec2-41 cells
(NY132), Sec4p peaks at the same density as in sec6-4 (NY779).
Sec2p partially cofractionates with Sec4p and Sncp in sec6-4 cells
and completely in sec2-41 cells.
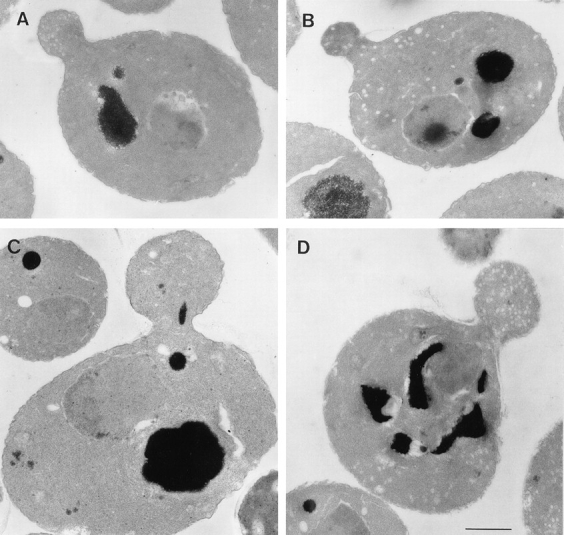
EM of sec2-78, wild-type, and sec6-4 cells shifted to 37°C for 10 min. Cells were synchronized with α factor, released from G1
arrest, and allowed to form small buds at 25°C, and then incubated at 37°C for 10 min. After this, cells were fixed and prepared for EM,
as described in Materials and Methods. (A) Representative sec2-78 cell (NY1529) at 25°C that shows some vesicle accumulation. (B)
Representative sec2-78 cell after 10 min at 37°C that accumulated vesicles both in bud and mother cell. In comparison, a typical wild-type cell (NY13; C) does not accumulate secretory vesicles, and in sec6-4 (NY17; D), vesicles accumulate mainly in the bud. Bar, 1 μm.
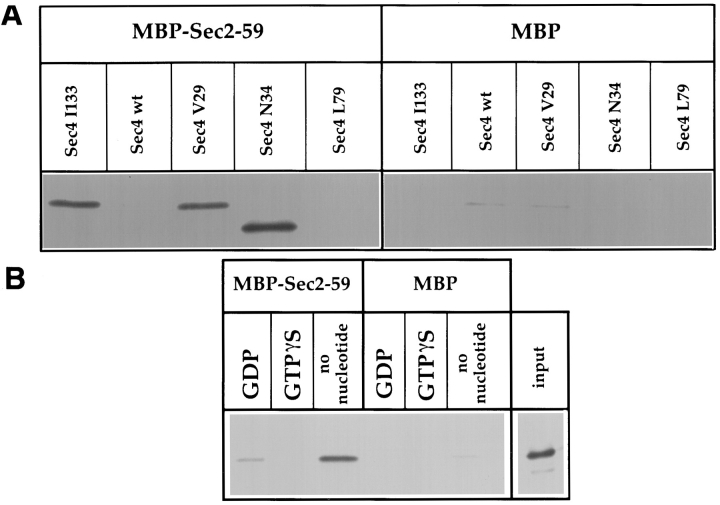
In vitro association
of Sec2p with Sec4p. The
binding assays were performed on amylose resin using recombinant MBP-Sec2-59 or MBP alone, which was
incubated with 35S-labeled
Sec4p produced in an in vitro
transcription-coupled translation reaction. (A) The
SEC4 mutations are indicated above each lane. (B)
Wild-type Sec4p was diluted
in buffer with EDTA before
being gel filtered and associated with nucleotide as indicated. Input was directly
loaded into the well and represents 40% of the 35S-labeled
protein used in each binding
reaction.
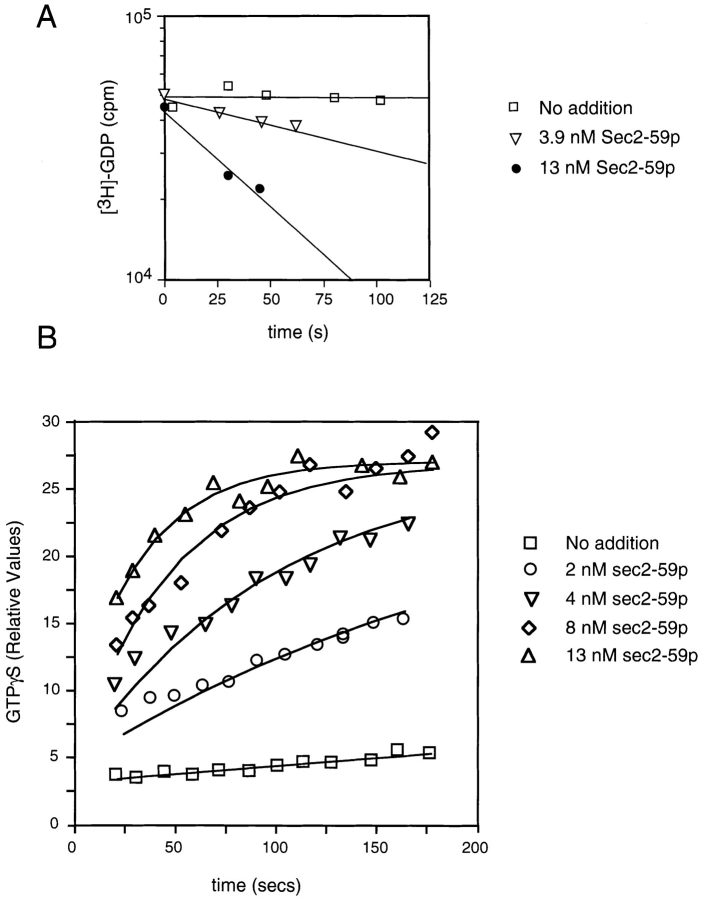
Sec2-59p–stimulated guanine nucleotide exchange on
Sec4p. (A) Aliquots containing 1 μM Sec4p-[3H]GDP were incubated with the indicated levels of Sec2-59p or buffer in the presence of excess unlabeled GDP. At the time intervals indicated,
Sec4p-bound radioactivity was determined by the filter-binding
assay. To retard the intrinsic off rate of Sec4p, this experiment
was performed at 14°C. (B) Aliquots containing 1 μM Sec4p were
incubated with Sec2-59p or buffer, as indicated in the presence of
[35S]GTPγS at 14°C. At the time intervals indicated, Sec4p-bound
radioactivity was determined by the filter-binding assay. For each
assay, the relative values correspond to the nanomoles of GTPγS
retained on the filter.

Sequence of Sec2p
and comparison with Rabin3
and expressed sequence tags.
Comparison of the Rab-binding
portion of the protein sequence
of Sec2p with Rabin3 and the
predicted protein sequence of
ESTs from human liver (H73617),
human brain (T09468/N24211),
and Caenorhabditis elegans
(D37628) homologues. The sequences were aligned using the
program MegAlign and visual
inspection. EST sequences, which
are incomplete cDNA sequences,
are shown only for the predicted
open reading frames corresponding to the SEC2 sequence.
Amino acid residues are numbered according to the protein
or DNA sequence. Gaps to improve the alignment are indicated by hyphens. Amino acids that are identical to the yeast sequence in at least two of the five sequences
are shaded black. Residues that are similar in class are shaded grey and are grouped according to acidic DE, basic HKR, hydrophobic
MPVWAFIL, and polar CGNQSTY.
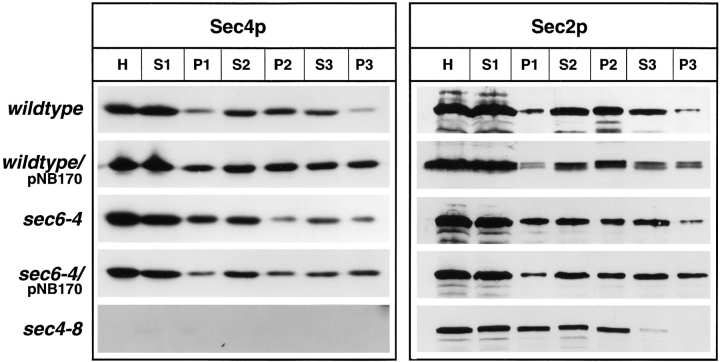
Differential centrifugation of lysates derived
from wild-type (NY10), wild-type overproducing Sec4p on
low copy (NY1535), sec6-4
(NY779), sec6-4 overproducing Sec4p (NY1536), and
sec4-8 (NY28). Cells were
grown overnight in YP dextrose and then shifted to
37°C. After 2 h at the restrictive temperature, cells were
harvested and lysates (S1)
were prepared as described
in Materials and Methods.
Differential centrifugation
and analysis of the resulting
fractions was performed as
described in Fig. 4. Western
blots were probed with
αSec4p polyclonal antibodies and iodinated protein A or affinity-purified αSec2p polyclonal antibodies detected with chemoluminescence. The amount of Sec2p in the P3 fractions parallels the amount of Sec4p. The increase of Sec4p in P3 of strains overproducing the
protein is correlated with an increase of Sec2p, and the decrease of Sec4p in P3 in sec4-8 cells is correlated with a decrease of Sec2p in
this fraction. The Sec4p blots for wild-type and sec6-4 cells are identical to those shown in Fig. 4 to allow for direct comparison.
Similar articles
-
Ypt32 recruits the Sec4p guanine nucleotide exchange factor, Sec2p, to secretory vesicles; evidence for a Rab cascade in yeast.J Cell Biol. 2002 Jun 10;157(6):1005-15. doi: 10.1083/jcb.200201003. Epub 2002 Jun 3. J Cell Biol. 2002. PMID: 12045183 Free PMC article.
-
The role of the COOH terminus of Sec2p in the transport of post-Golgi vesicles.J Cell Biol. 2000 Apr 3;149(1):95-110. doi: 10.1083/jcb.149.1.95. J Cell Biol. 2000. PMID: 10747090 Free PMC article.
-
Interactions of nucleotide release factor Dss4p with Sec4p in the post-Golgi secretory pathway of yeast.J Biol Chem. 1997 Jul 18;272(29):18281-9. doi: 10.1074/jbc.272.29.18281. J Biol Chem. 1997. PMID: 9218467
-
The cycle of SEC4 function in vesicular transport.Ciba Found Symp. 1993;176:218-28; discussion 229-32. Ciba Found Symp. 1993. PMID: 8299422 Review.
-
Small GTP-binding proteins and their role in transport.Curr Opin Cell Biol. 1991 Aug;3(4):626-33. doi: 10.1016/0955-0674(91)90033-u. Curr Opin Cell Biol. 1991. PMID: 1663370 Review. No abstract available.
Cited by
-
Structural basis for recognition of the Sec4 Rab GTPase by its effector, the Lgl/tomosyn homologue, Sro7.Mol Biol Cell. 2015 Sep 15;26(18):3289-300. doi: 10.1091/mbc.E15-04-0228. Epub 2015 Jul 22. Mol Biol Cell. 2015. PMID: 26202462 Free PMC article.
-
Exocyst sec5 regulates exocytosis of newcomer insulin granules underlying biphasic insulin secretion.PLoS One. 2013 Jul 2;8(7):e67561. doi: 10.1371/journal.pone.0067561. Print 2013. PLoS One. 2013. PMID: 23844030 Free PMC article.
-
Two distinct regions in a yeast myosin-V tail domain are required for the movement of different cargoes.J Cell Biol. 2000 Aug 7;150(3):513-26. doi: 10.1083/jcb.150.3.513. J Cell Biol. 2000. PMID: 10931864 Free PMC article.
-
Family-wide characterization of the DENN domain Rab GDP-GTP exchange factors.J Cell Biol. 2010 Oct 18;191(2):367-81. doi: 10.1083/jcb.201008051. Epub 2010 Oct 11. J Cell Biol. 2010. PMID: 20937701 Free PMC article.
-
The exocyst complex in exocytosis and cell migration.Protoplasma. 2012 Jul;249(3):587-97. doi: 10.1007/s00709-011-0330-1. Epub 2011 Oct 14. Protoplasma. 2012. PMID: 21997494 Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
