

doi: 10.1073/pnas.94.12.6084.
Transient aggregates in protein folding are easily mistaken for folding intermediates
Affiliations
- PMID: 9177173
- PMCID: PMC21005
- DOI: 10.1073/pnas.94.12.6084
Item in Clipboard
Transient aggregates in protein folding are easily mistaken for folding intermediates
Proc Natl Acad Sci U S A.
.
Abstract
It has been questioned recently whether populated intermediates are important for the protein folding process or are artefacts trapped in nonproductive pathways. We report here that the rapidly formed intermediate of the spliceosomal protein U1A is an off-pathway artefact caused by transient aggregation of denatured protein under native conditions. Transient aggregates are easily mistaken for structured monomers and could be a general problem in time-resolved folding studies.
Figures
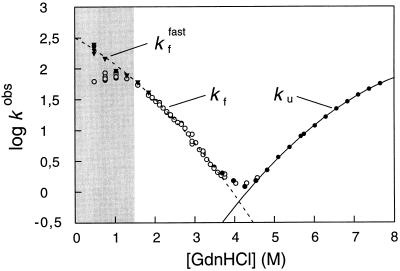
Gdn⋅HCl dependence of the rate constants for folding and unfolding of U1A. The rate constants are in units of s−1. The left arm of the V-shaped plot shows the refolding rate constant (○, kf, [U1A] = 3.1 μM; and ▾, kffast, [U1A] = 1 μM) following 1:10 dilution (stopped-flow) of denatured U1A (in 5.1 M Gdn⋅HCl) into lower [Gdn⋅HCl], and the right arm shows the unfolding rate constant (•, ku, [U1A] = 3.1 μM) upon 1:10 mixing of native protein (in water) into high [Gdn⋅HCl]. The curves are polynomial fits which precisely obey Eq. 1 and, hence, represent two-state folding directly from the denatured state—i.e., log kf = log ku − log KD–N. The deviation from two-state folding observed at low [Gdn⋅HCl] (○) is found also for other proteins and is usually believed to result from accumulation of an intermediate. With U1A, the deviation is caused by transient aggregation of denatured protein under refolding conditions. At low protein concentrations the denatured protein remains monomeric during the refolding process and the rate constant (kffast) follows Eq. 1, but at higher protein concentrations the denatured protein aggregates in the dead-time of the stopped-flow instrument, giving rise to a retardation of the refolding rate. Conditions where aggregation occurs are marked gray.
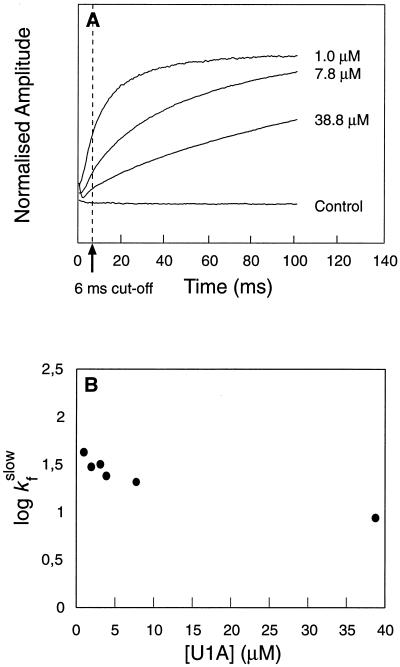
(A) Time course for refolding of U1A at different protein concentrations. Final [Gdn⋅HCl] = 0.46 M. At moderate to high protein concentrations (>5 μM), the time course is dominated by the slow phase, but at low protein concentrations folding occurs mainly by the fast reaction. (B) The rate constant of the slow phase decreases slightly with increasing protein concentration, whereas the fast reaction appears independent of protein concentration. The negative concentration dependence of the slow phase is inconsistent with formation of aggregates, since this process would become faster at high protein concentrations. Hence, it is likely that the slow phase represents a dissociation process—i.e., folding from an aggregate. Data from the first 6 ms were excluded from the fits. Control experiments were conducted with free tryptophan and with U1A contained in the dilution buffer.
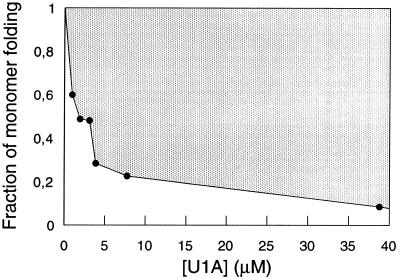
Fraction of monomer folding at different concentrations of U1A, expressed as the ratio of the amplitudes of the fast and slow refolding phase (compare Fig. 2A). In fits where [U1A] ≥ 3.1 μM the rate constant for the fast phase was locked to 200 s−1. Since refolding is usually monitored at relatively high concentrations of protein, the proportion of monomer folding may be very small and undetected. For example, standard stopped-flow (≈10 μM), stopped-flow CD (10–50 μM), and quench-flow NMR (>100 μM). Hence, tests of concentration dependence in these regions may not reveal aggregation artefacts.
Similar articles
-
Transient aggregation and stable dimerization induced by introducing an Alzheimer sequence into a water-soluble protein.Biochemistry. 2004 Oct 19;43(41):12964-78. doi: 10.1021/bi048509k. Biochemistry. 2004. PMID: 15476390
-
High-energy channeling in protein folding.Biochemistry. 1997 Jun 24;36(25):7633-7. doi: 10.1021/bi970210x. Biochemistry. 1997. PMID: 9201903
-
Formation of short-lived protein aggregates directly from the coil in two-state folding.Biochemistry. 1999 Oct 5;38(40):13006-12. doi: 10.1021/bi9909997. Biochemistry. 1999. PMID: 10529170
-
Spliceosomal UsnRNP biogenesis, structure and function.Curr Opin Cell Biol. 2001 Jun;13(3):290-301. doi: 10.1016/s0955-0674(00)00211-8. Curr Opin Cell Biol. 2001. PMID: 11343899 Review.
-
Architecture of the spliceosome.Biochemistry. 2012 Apr 24;51(16):3321-33. doi: 10.1021/bi201215r. Epub 2012 Apr 10. Biochemistry. 2012. PMID: 22471593 Review.
Cited by
-
The sequences of small proteins are not extensively optimized for rapid folding by natural selection.Proc Natl Acad Sci U S A. 1998 Apr 28;95(9):4982-6. doi: 10.1073/pnas.95.9.4982. Proc Natl Acad Sci U S A. 1998. PMID: 9560214 Free PMC article.
-
A protein folding pathway with multiple folding intermediates at atomic resolution.Proc Natl Acad Sci U S A. 2005 Apr 5;102(14):5026-31. doi: 10.1073/pnas.0501372102. Epub 2005 Mar 25. Proc Natl Acad Sci U S A. 2005. PMID: 15793003 Free PMC article.
-
Folding of human superoxide dismutase: disulfide reduction prevents dimerization and produces marginally stable monomers.Proc Natl Acad Sci U S A. 2004 Nov 9;101(45):15893-8. doi: 10.1073/pnas.0403979101. Epub 2004 Nov 2. Proc Natl Acad Sci U S A. 2004. PMID: 15522970 Free PMC article.
-
Competition of individual domain folding with inter-domain interaction in WW domain engineered repeat proteins.Phys Chem Chem Phys. 2019 Nov 13;21(44):24393-24405. doi: 10.1039/c8cp07775d. Phys Chem Chem Phys. 2019. PMID: 31663524 Free PMC article.
-
Observation of strange kinetics in protein folding.Proc Natl Acad Sci U S A. 1999 May 25;96(11):6031-6. doi: 10.1073/pnas.96.11.6031. Proc Natl Acad Sci U S A. 1999. PMID: 10339536 Free PMC article.
References
-
- Udgaonkar J B, Baldwin R L. Nature (London) 1988;335:700–704. - PubMed
-
- Matouschek A, Kellis J T, Serrano L, Bycroft M, Fersht A R. Nature (London) 1990;346:440–445. - PubMed
-
- Kim P S, Baldwin R L. Annu Rev Biochem. 1990;59:631–660. - PubMed
-
- Radford S E, Dobson C M, Evans P A. Nature (London) 1992;358:302–307. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
