

Second messengers, trafficking-related proteins, and amino acid residues that contribute to the functional regulation of the rat brain GABA transporter GAT1
- PMID: 9096133
- PMCID: PMC6573650
- DOI: 10.1523/JNEUROSCI.17-09-02967.1997
Second messengers, trafficking-related proteins, and amino acid residues that contribute to the functional regulation of the rat brain GABA transporter GAT1
Abstract
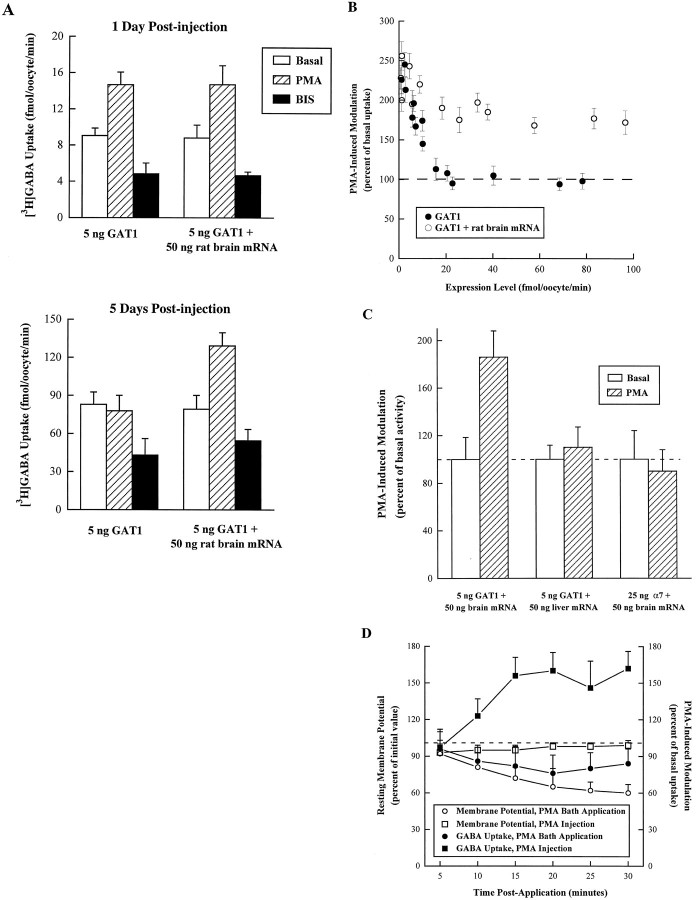
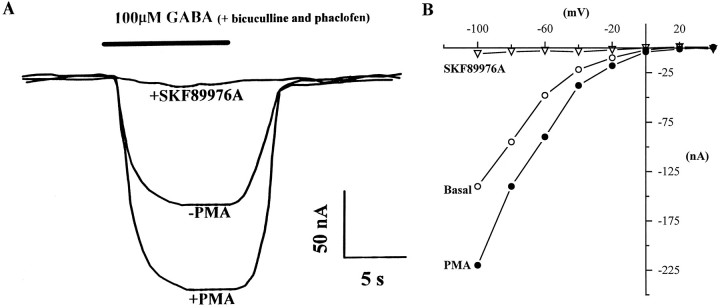
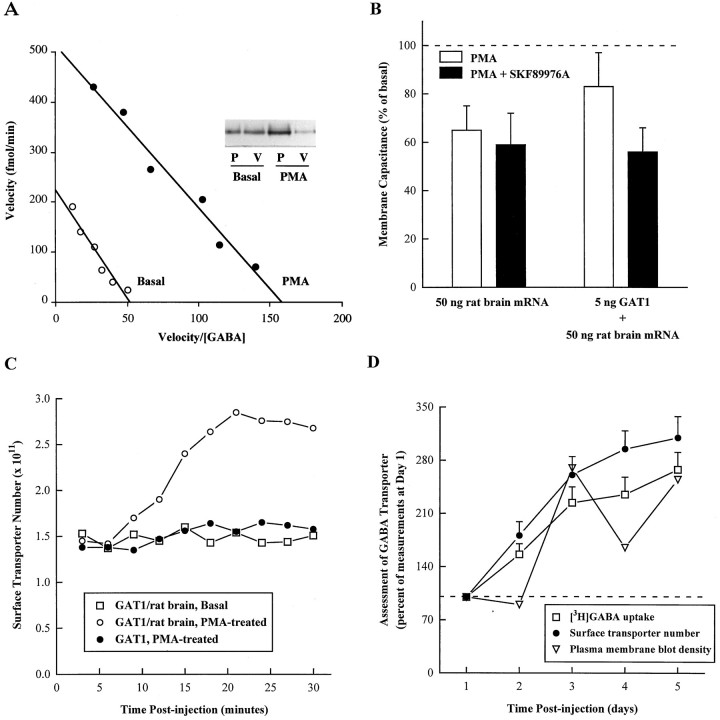
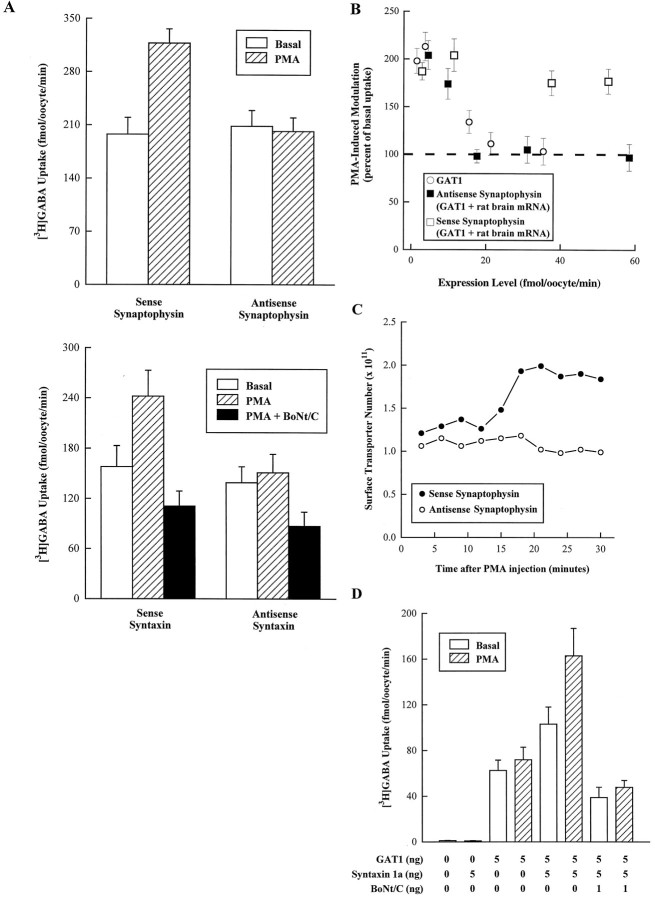
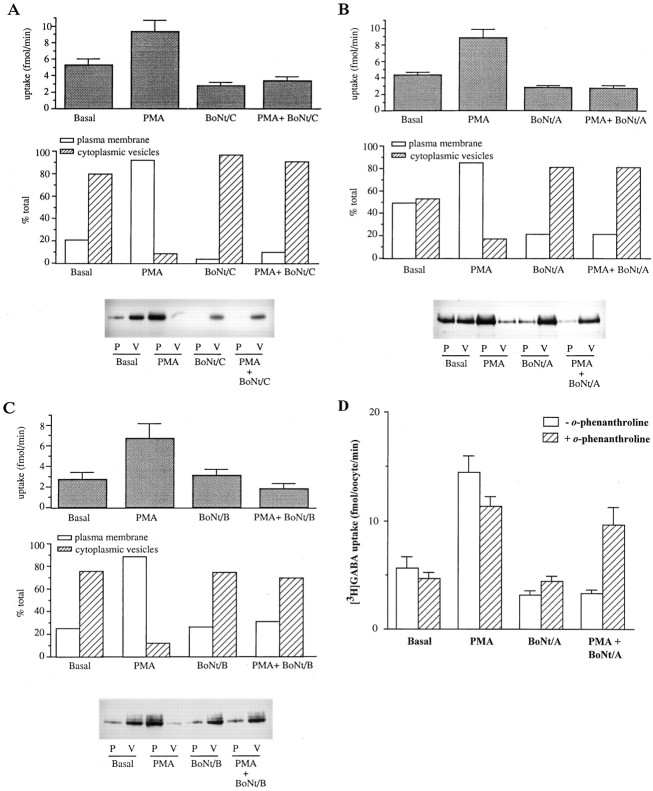
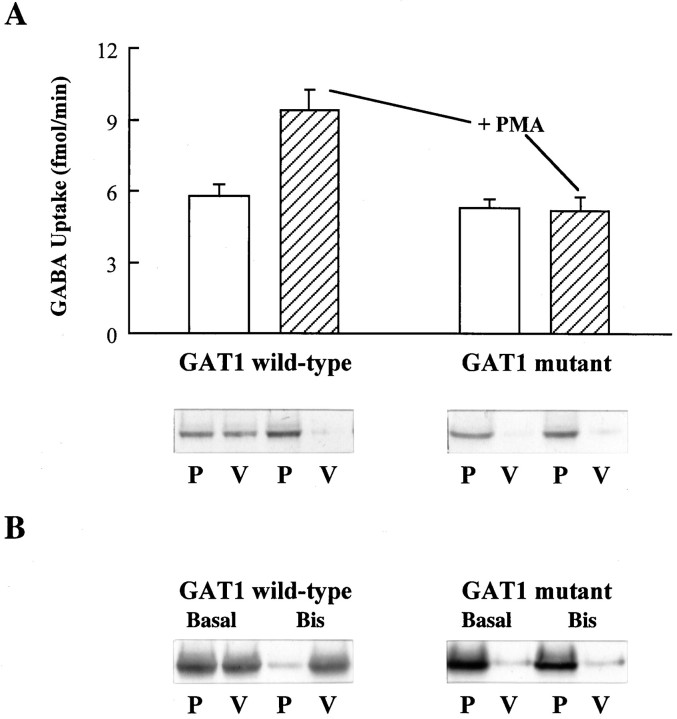
Recent evidence indicates that several members of the Na+-coupled transporter family are regulated, and this regulation in part occurs by redistribution of transporters between intracellular locations and the plasma membrane. We elucidate components of this process for both wild-type and mutant GABA transporters (GAT1) expressed in Xenopus oocytes using a combination of uptake assays, immunoblots, and electrophysiological measurements of membrane capacitance, transport-associated currents, and GAT1-specific charge movements. At low GAT1 expression levels, activators of protein kinase C (PKC) induce redistribution of GAT1 from intracellular vesicles to the plasma membrane; at higher GAT1 expression levels, activators of PKC fail to induce this redistribution. However, coinjection of total rat brain mRNA with GAT1 permits PKC-mediated modulation at high transporter expression levels. This effect of brain mRNA on modulation is mimicked by coinjection of syntaxin 1a mRNA and is eliminated by injecting synaptophysin or syntaxin antisense oligonucleotides. Additionally, botulinum toxins, which inactivate proteins involved in vesicle release and recycling, reduce basal GAT1 expression and prevent PKC-induced translocation. Mutant GAT1 proteins, in which most or all of a leucine heptad repeat sequence was removed, display altered basal distribution and lack susceptibility to modulation by PKC, delineating one region of GAT1 necessary for its targeting. Thus, functional regulation of GAT1 in oocytes occurs via components common to transporters and to trafficking in both neural and non-neural cells, and suggests a relationship between factors that control neurotransmitter secretion and the components necessary for neurotransmitter uptake.
Figures
Similar articles
-
Protein kinase C modulates the activity of a cloned gamma-aminobutyric acid transporter expressed in Xenopus oocytes via regulated subcellular redistribution of the transporter.J Biol Chem. 1994 May 20;269(20):14759-67. J Biol Chem. 1994. PMID: 8182081
-
Syntaxin 1A up-regulates GABA transporter expression by subcellular redistribution.Mol Membr Biol. 2001 Jan-Mar;18(1):39-44. Mol Membr Biol. 2001. PMID: 11396610
-
Syntaxin 1A inhibits GABA flux, efflux, and exchange mediated by the rat brain GABA transporter GAT1.Mol Pharmacol. 2003 Oct;64(4):905-13. doi: 10.1124/mol.64.4.905. Mol Pharmacol. 2003. PMID: 14500747
-
Structure, function and brain localization of neurotransmitter transporters.J Exp Biol. 1994 Nov;196:283-95. doi: 10.1242/jeb.196.1.283. J Exp Biol. 1994. PMID: 7823028 Review.
-
TRP Channel Trafficking.In: Liedtke WB, Heller S, editors. TRP Ion Channel Function in Sensory Transduction and Cellular Signaling Cascades. Boca Raton (FL): CRC Press/Taylor & Francis; 2007. Chapter 23. In: Liedtke WB, Heller S, editors. TRP Ion Channel Function in Sensory Transduction and Cellular Signaling Cascades. Boca Raton (FL): CRC Press/Taylor & Francis; 2007. Chapter 23. PMID: 21204515 Free Books & Documents. Review.
Cited by
-
Sec24- and ARFGAP1-dependent trafficking of GABA transporter-1 is a prerequisite for correct axonal targeting.J Neurosci. 2008 Nov 19;28(47):12453-64. doi: 10.1523/JNEUROSCI.3451-08.2008. J Neurosci. 2008. PMID: 19020038 Free PMC article.
-
Multiple signaling pathways regulate cell surface expression and activity of the excitatory amino acid carrier 1 subtype of Glu transporter in C6 glioma.J Neurosci. 1998 Apr 1;18(7):2475-85. doi: 10.1523/JNEUROSCI.18-07-02475.1998. J Neurosci. 1998. PMID: 9502808 Free PMC article.
-
Inhibition of Activity of GABA Transporter GAT1 by δ-Opioid Receptor.Evid Based Complement Alternat Med. 2012;2012:818451. doi: 10.1155/2012/818451. Epub 2012 Dec 25. Evid Based Complement Alternat Med. 2012. PMID: 23365600 Free PMC article.
-
Anion currents and predicted glutamate flux through a neuronal glutamate transporter.J Neurosci. 1998 Sep 15;18(18):7099-110. doi: 10.1523/JNEUROSCI.18-18-07099.1998. J Neurosci. 1998. PMID: 9736633 Free PMC article.
-
Substrates regulate gamma-aminobutyric acid transporters in a syntaxin 1A-dependent manner.Proc Natl Acad Sci U S A. 2002 Apr 16;99(8):5686-91. doi: 10.1073/pnas.082712899. Proc Natl Acad Sci U S A. 2002. PMID: 11960023 Free PMC article.
References
-
- Ahn J, Mundigl O, Muth TR, Rudnick G, Caplan MJ. Polarized expression of GABA transporters in Madin-Darby canine kidney cells and cultured hippocampal neurons. J Biol Chem. 1996;271:6917–6924. - PubMed
-
- Alder J, Lu B, Valtorta F, Greengard P, Poo M. Calcium-dependent transmitter secretion reconstituted in Xenopus oocytes: requirement for synaptophysin. Science. 1992;257:657–661. - PubMed
-
- Asano T, Takata K, Katagiri H, Tsukuda K, Lin J-L, Ishihara H, Inukai K, Hirano H, Yazaki Y, Oka Y. Domains responsible for the differential targeting of glucose transporter isoforms. J Biol Chem. 1992;267:19636–19641. - PubMed
-
- Bennett MK, Calakos N, Scheller RH. Syntaxin: a synaptic protein implicated in docking of synaptic vesicles at presynaptic active zones. Science. 1992;257:255–259. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources