

Structure-function analyses reveal key molecular determinants of HIV-1 CRF01_AE resistance to the entry inhibitor temsavir
- PMID: 37872202
- PMCID: PMC10593844
- DOI: 10.1038/s41467-023-42500-2
Structure-function analyses reveal key molecular determinants of HIV-1 CRF01_AE resistance to the entry inhibitor temsavir
Abstract
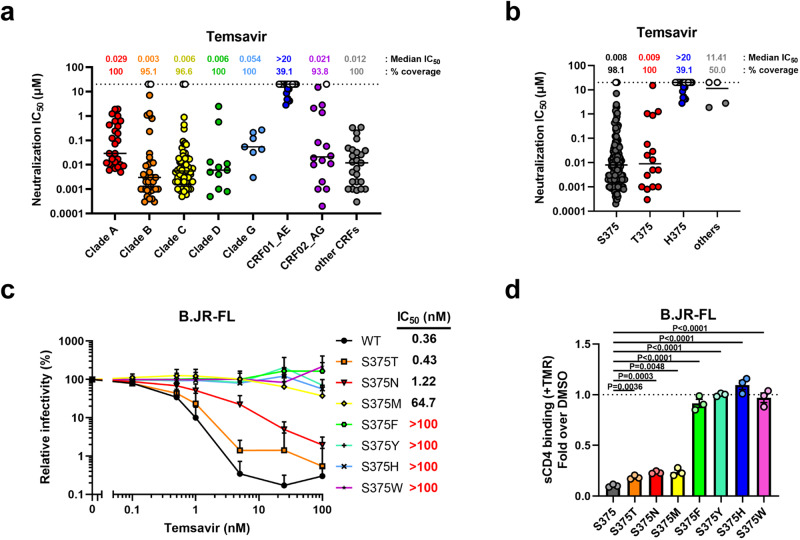
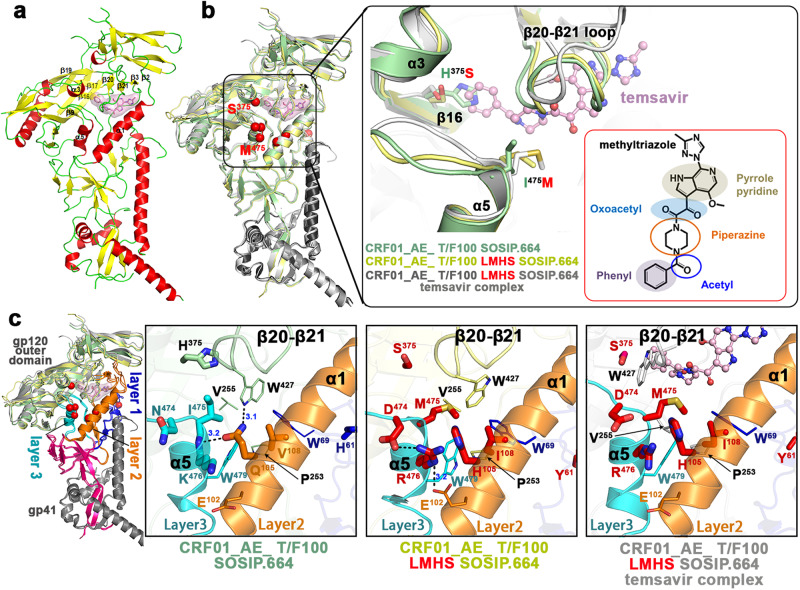
The HIV-1 entry inhibitor temsavir prevents the viral receptor CD4 (cluster of differentiation 4) from interacting with the envelope glycoprotein (Env) and blocks its conformational changes. To do this, temsavir relies on the presence of a residue with small side chain at position 375 in Env and is unable to neutralize viral strains like CRF01_AE carrying His375. Here we investigate the mechanism of temsavir resistance and show that residue 375 is not the sole determinant of resistance. At least six additional residues within the gp120 inner domain layers, including five distant from the drug-binding pocket, contribute to resistance. A detailed structure-function analysis using engineered viruses and soluble trimer variants reveals that the molecular basis of resistance is mediated by crosstalk between His375 and the inner domain layers. Furthermore, our data confirm that temsavir can adjust its binding mode to accommodate changes in Env conformation, a property that likely contributes to its broad antiviral activity.
© 2023. Springer Nature Limited.
Conflict of interest statement
J.P., A.F., and M.P., are inventors on U.S. patent application No. 17/762,333 related to compositions and methods based on HIV gp120 LMHS mutants, in which there are no restrictions on the publication of this manuscript data. The remaining authors declare no competing interests. The views expressed in this manuscript are those of the authors and do not reflect the official policy or position of the Uniformed Services University, US Army, the Department of Defense, or the US Government.
Figures




Update of
-
Structure-function Analyses Reveal Key Molecular Determinants of HIV-1 CRF01_AE Resistance to the Entry Inhibitor Temsavir.bioRxiv [Preprint]. 2023 Apr 17:2023.04.17.537181. doi: 10.1101/2023.04.17.537181. bioRxiv. 2023. Update in: Nat Commun. 2023 Oct 23;14(1):6710. doi: 10.1038/s41467-023-42500-2. PMID: 37131729 Free PMC article. Updated. Preprint.
Similar articles
-
Structure-function Analyses Reveal Key Molecular Determinants of HIV-1 CRF01_AE Resistance to the Entry Inhibitor Temsavir.bioRxiv [Preprint]. 2023 Apr 17:2023.04.17.537181. doi: 10.1101/2023.04.17.537181. bioRxiv. 2023. Update in: Nat Commun. 2023 Oct 23;14(1):6710. doi: 10.1038/s41467-023-42500-2. PMID: 37131729 Free PMC article. Updated. Preprint.
-
Histidine 375 Modulates CD4 Binding in HIV-1 CRF01_AE Envelope Glycoproteins.J Virol. 2017 Jan 31;91(4):e02151-16. doi: 10.1128/JVI.02151-16. Print 2017 Feb 15. J Virol. 2017. PMID: 27928014 Free PMC article.
-
Activation and Inactivation of Primary Human Immunodeficiency Virus Envelope Glycoprotein Trimers by CD4-Mimetic Compounds.J Virol. 2017 Jan 18;91(3):e01880-16. doi: 10.1128/JVI.01880-16. Print 2017 Feb 1. J Virol. 2017. PMID: 27881646 Free PMC article.
-
Small Molecule HIV-1 Attachment Inhibitors: Discovery, Mode of Action and Structural Basis of Inhibition.Viruses. 2021 May 6;13(5):843. doi: 10.3390/v13050843. Viruses. 2021. PMID: 34066522 Free PMC article. Review.
-
Small-molecule HIV-1 gp120 inhibitors to prevent HIV-1 entry: an emerging opportunity for drug development.Curr Opin Investig Drugs. 2006 Aug;7(8):721-6. Curr Opin Investig Drugs. 2006. PMID: 16955683 Review.
Cited by
-
Conformational dynamics of the HIV-1 envelope glycoprotein from CRF01_AE is associated with susceptibility to antibody-dependent cellular cytotoxicity.bioRxiv [Preprint]. 2024 Aug 22:2024.08.22.609179. doi: 10.1101/2024.08.22.609179. bioRxiv. 2024. PMID: 39229074 Free PMC article. Preprint.
-
Temsavir Modulates HIV-1 Envelope Conformation by Decreasing Its Proteolytic Cleavage.Viruses. 2023 May 18;15(5):1189. doi: 10.3390/v15051189. Viruses. 2023. PMID: 37243275 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
