

SEC14-like condensate phase transitions at plasma membranes regulate root growth in Arabidopsis
- PMID: 37721949
- PMCID: PMC10538751
- DOI: 10.1371/journal.pbio.3002305
SEC14-like condensate phase transitions at plasma membranes regulate root growth in Arabidopsis
Abstract
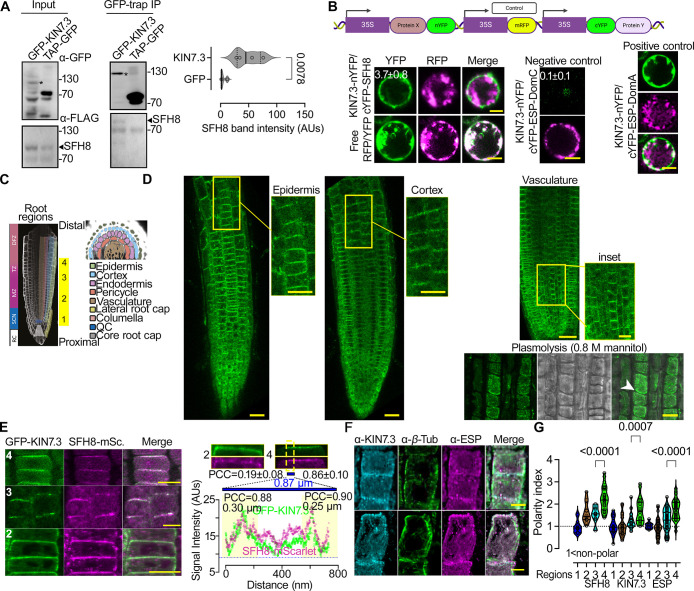
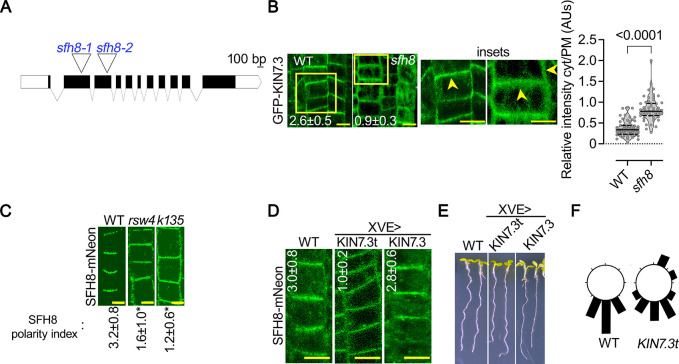
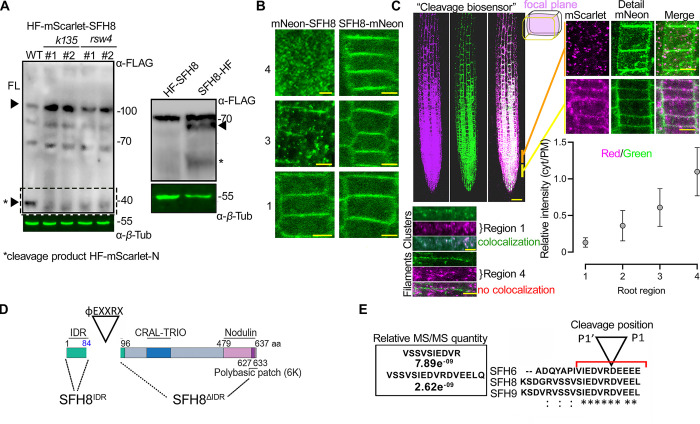
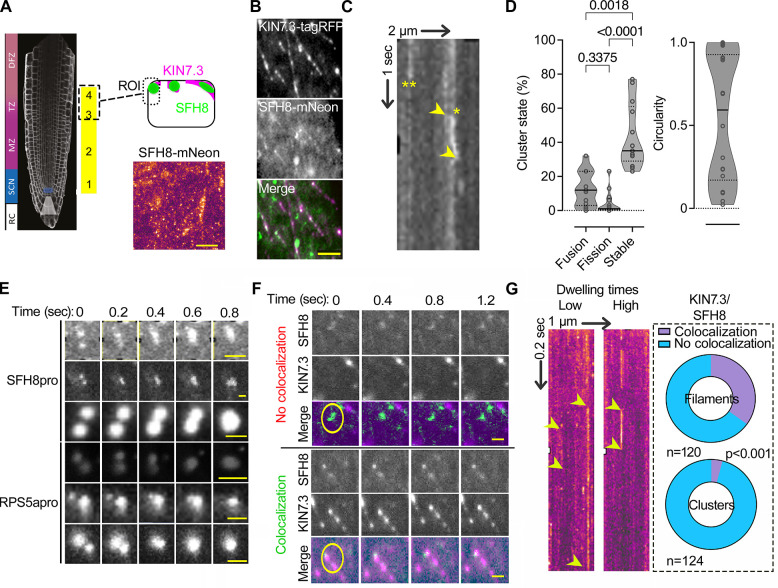
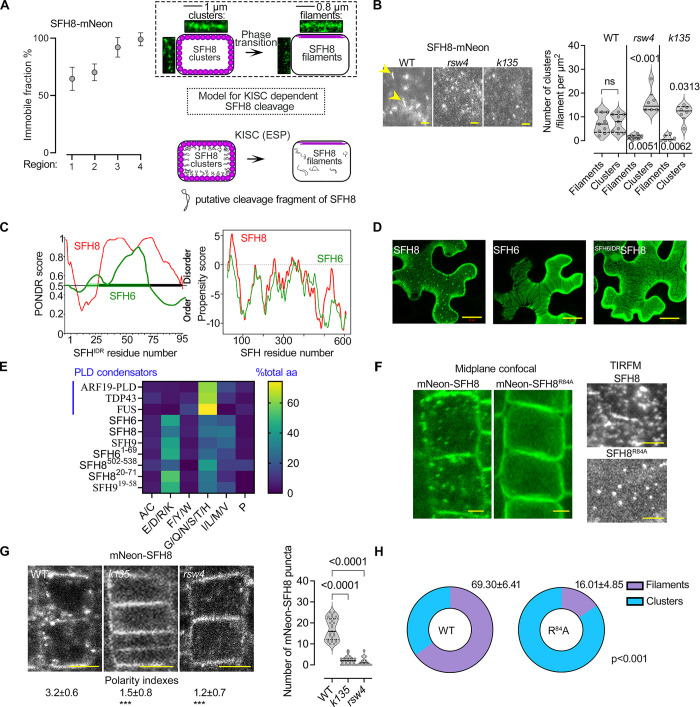
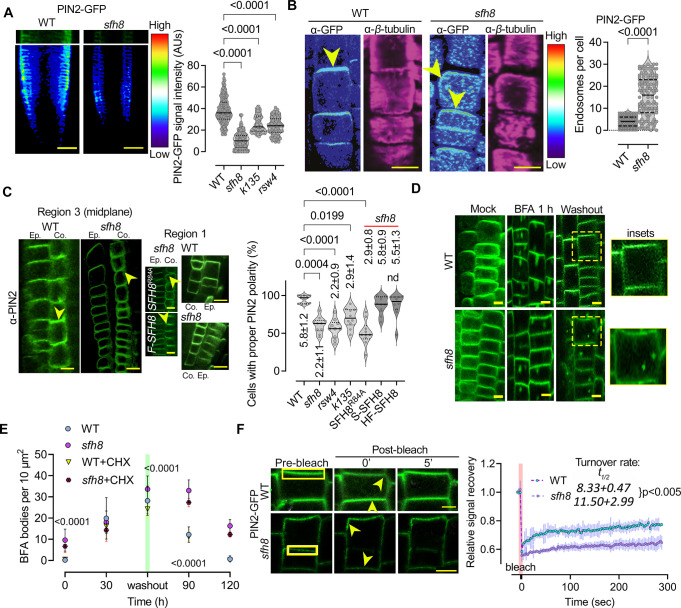
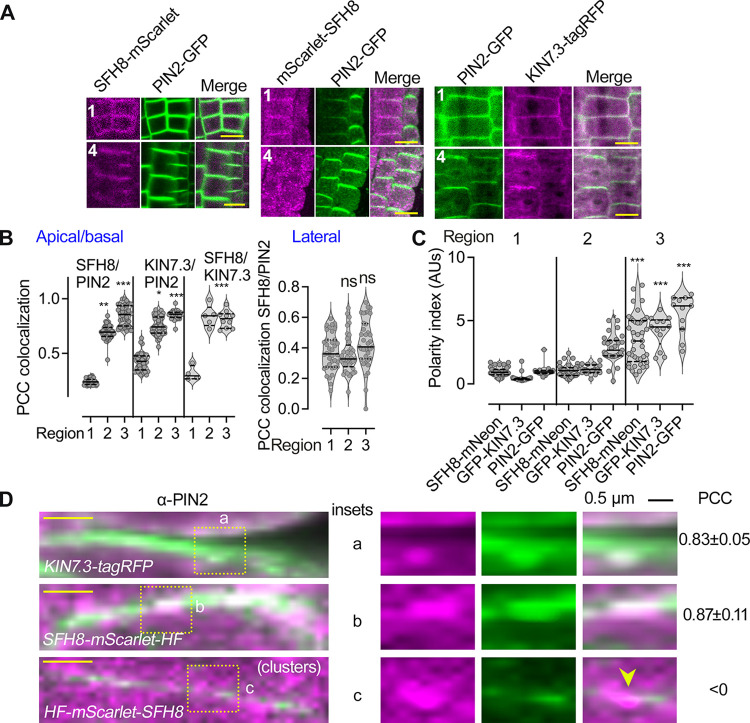
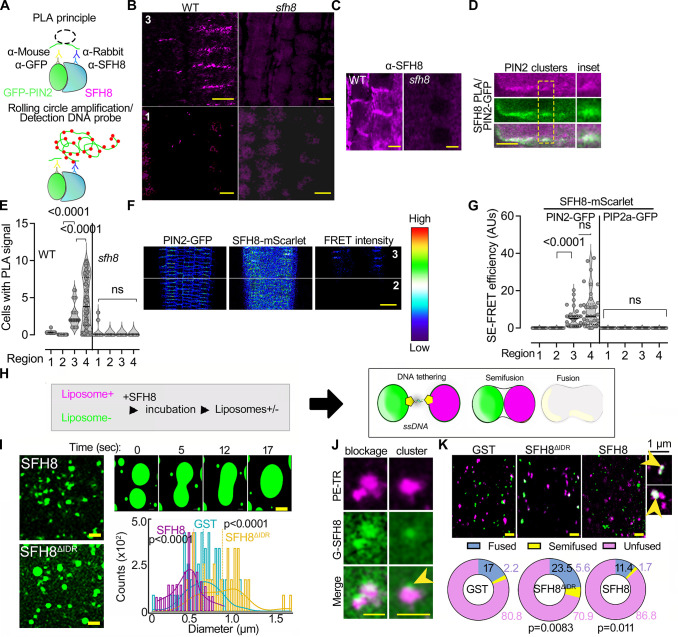
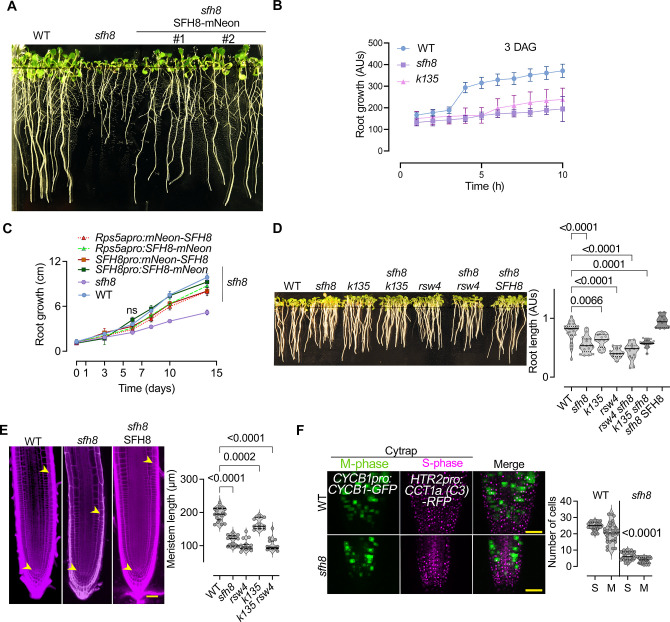
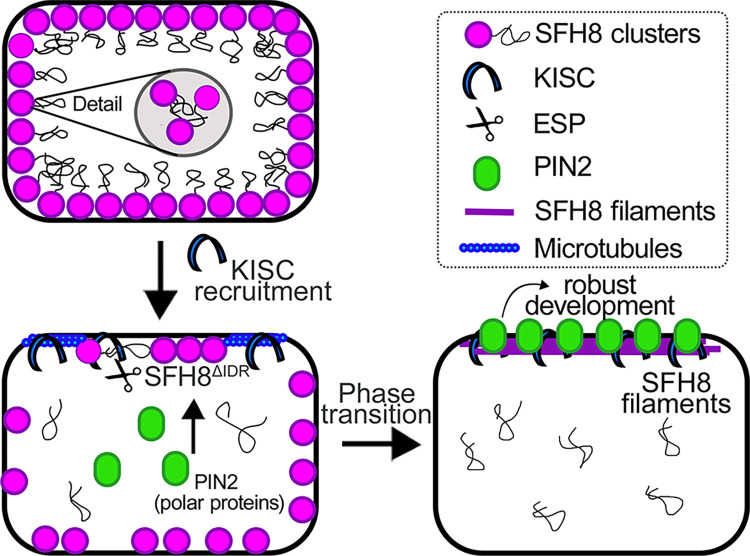
Protein function can be modulated by phase transitions in their material properties, which can range from liquid- to solid-like; yet, the mechanisms that drive these transitions and whether they are important for physiology are still unknown. In the model plant Arabidopsis, we show that developmental robustness is reinforced by phase transitions of the plasma membrane-bound lipid-binding protein SEC14-like. Using imaging, genetics, and in vitro reconstitution experiments, we show that SEC14-like undergoes liquid-like phase separation in the root stem cells. Outside the stem cell niche, SEC14-like associates with the caspase-like protease separase and conserved microtubule motors at unique polar plasma membrane interfaces. In these interfaces, SEC14-like undergoes processing by separase, which promotes its liquid-to-solid transition. This transition is important for root development, as lines expressing an uncleavable SEC14-like variant or mutants of separase and associated microtubule motors show similar developmental phenotypes. Furthermore, the processed and solidified but not the liquid form of SEC14-like interacts with and regulates the polarity of the auxin efflux carrier PINFORMED2. This work demonstrates that robust development can involve liquid-to-solid transitions mediated by proteolysis at unique plasma membrane interfaces.
Copyright: © 2023 Liu et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
The caspase-related protease separase (extra spindle poles) regulates cell polarity and cytokinesis in Arabidopsis.Plant Cell. 2013 Jun;25(6):2171-86. doi: 10.1105/tpc.113.113043. Epub 2013 Jun 28. Plant Cell. 2013. PMID: 23898031 Free PMC article.
-
SEC14-GOLD protein PATELLIN2 binds IRON-REGULATED TRANSPORTER1 linking root iron uptake to vitamin E.Plant Physiol. 2023 May 2;192(1):504-526. doi: 10.1093/plphys/kiac563. Plant Physiol. 2023. PMID: 36493393 Free PMC article.
-
Phylogenetic analysis of plant multi-domain SEC14-like phosphatidylinositol transfer proteins and structure-function properties of PATELLIN2.Plant Mol Biol. 2020 Dec;104(6):665-678. doi: 10.1007/s11103-020-01067-y. Epub 2020 Sep 11. Plant Mol Biol. 2020. PMID: 32915352 Free PMC article.
-
Role of SEC14-like phosphatidylinositol transfer proteins in membrane identity and dynamics.Front Plant Sci. 2023 May 15;14:1181031. doi: 10.3389/fpls.2023.1181031. eCollection 2023. Front Plant Sci. 2023. PMID: 37255567 Free PMC article. Review.
-
Surprising roles for phospholipid binding proteins revealed by high throughput genetics.Biochem Cell Biol. 2010 Aug;88(4):565-74. doi: 10.1139/O09-171. Biochem Cell Biol. 2010. PMID: 20651827 Review.
Cited by
-
Basic design of artificial membrane-less organelles using condensation-prone proteins in plant cells.Commun Biol. 2024 Oct 26;7(1):1396. doi: 10.1038/s42003-024-07102-8. Commun Biol. 2024. PMID: 39462114 Free PMC article.
-
Guidelines for naming and studying plasma membrane domains in plants.Nat Plants. 2024 Aug;10(8):1172-1183. doi: 10.1038/s41477-024-01742-8. Epub 2024 Aug 12. Nat Plants. 2024. PMID: 39134664 Review.
-
Seed longevity is controlled by metacaspases.Nat Commun. 2024 Aug 8;15(1):6748. doi: 10.1038/s41467-024-50848-2. Nat Commun. 2024. PMID: 39117606 Free PMC article.
-
Stress-related biomolecular condensates in plants.Plant Cell. 2023 Sep 1;35(9):3187-3204. doi: 10.1093/plcell/koad127. Plant Cell. 2023. PMID: 37162152 Free PMC article.
-
Protein phase separation in plant membrane biology: more than just a compartmentalization strategy.Plant Cell. 2023 Sep 1;35(9):3162-3172. doi: 10.1093/plcell/koad177. Plant Cell. 2023. PMID: 37352127 Free PMC article.
References
Grants and funding
LinkOut - more resources
Full Text Sources
