

The material properties of a bacterial-derived biomolecular condensate tune biological function in natural and synthetic systems
- PMID: 36163138
- PMCID: PMC9512792
- DOI: 10.1038/s41467-022-33221-z
The material properties of a bacterial-derived biomolecular condensate tune biological function in natural and synthetic systems
Abstract
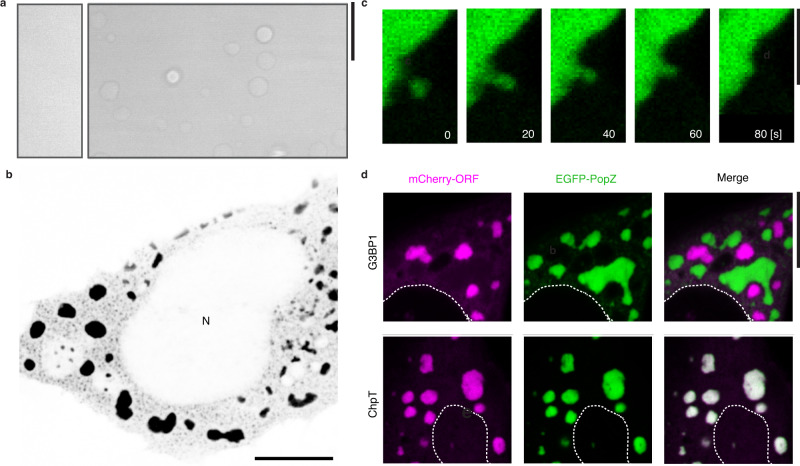
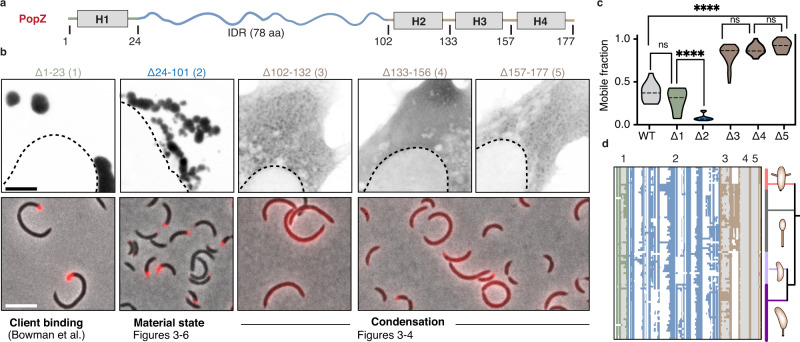
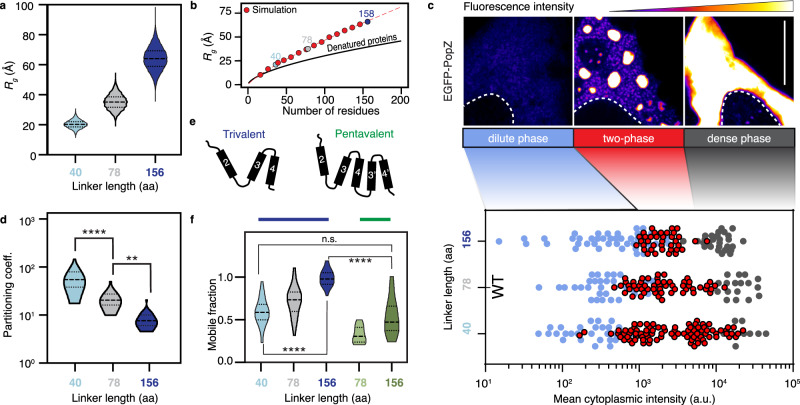
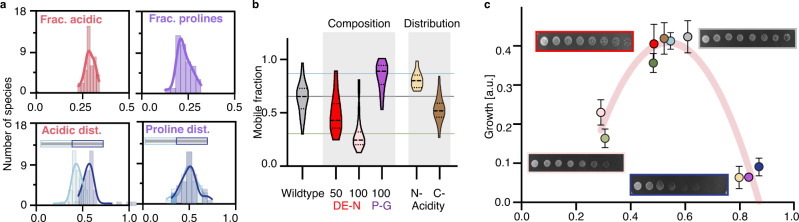
Intracellular phase separation is emerging as a universal principle for organizing biochemical reactions in time and space. It remains incompletely resolved how biological function is encoded in these assemblies and whether this depends on their material state. The conserved intrinsically disordered protein PopZ forms condensates at the poles of the bacterium Caulobacter crescentus, which in turn orchestrate cell-cycle regulating signaling cascades. Here we show that the material properties of these condensates are determined by a balance between attractive and repulsive forces mediated by a helical oligomerization domain and an expanded disordered region, respectively. A series of PopZ mutants disrupting this balance results in condensates that span the material properties spectrum, from liquid to solid. A narrow range of condensate material properties supports proper cell division, linking emergent properties to organismal fitness. We use these insights to repurpose PopZ as a modular platform for generating tunable synthetic condensates in human cells.
© 2022. The Author(s).
Conflict of interest statement
A.S.H. is a scientific consultant with Dewpoint Therapeutics. A.D.G. has served as a consultant for Aquinnah Pharmaceuticals, Prevail Therapeutics and Third Rock Ventures and is a scientific founder of Maze Therapeutics. L.S. is on the Board of Directors of Pacific Biosciences, Inc and 5Metis, Inc. Chan Zuckerberg Biohub, Inc. and The Board Of Trustees Of The Leland Stanford Junior University filed a patent application that describes the PopTag technology, parts of which are introduced and described in this work, on behalf of inventors K.L., S.B, A.D.G, and L.S. (PCT/ WO2021113598A3). V.L, D.S., E.S., A.B., M.J., D.D., A.D., and E.V. declare no competing interests.
Figures



Similar articles
-
Caulobacter PopZ forms an intrinsically disordered hub in organizing bacterial cell poles.Proc Natl Acad Sci U S A. 2016 Nov 1;113(44):12490-12495. doi: 10.1073/pnas.1602380113. Epub 2016 Oct 18. Proc Natl Acad Sci U S A. 2016. PMID: 27791060 Free PMC article.
-
A Localized Complex of Two Protein Oligomers Controls the Orientation of Cell Polarity.mBio. 2017 Feb 28;8(1):e02238-16. doi: 10.1128/mBio.02238-16. mBio. 2017. PMID: 28246363 Free PMC article.
-
Sequence determinants of in cell condensate morphology, dynamics, and oligomerization as measured by number and brightness analysis.Cell Commun Signal. 2021 Jun 5;19(1):65. doi: 10.1186/s12964-021-00744-9. Cell Commun Signal. 2021. PMID: 34090478 Free PMC article.
-
Conformational Dynamics of Intrinsically Disordered Proteins Regulate Biomolecular Condensate Chemistry.Chem Rev. 2022 Mar 23;122(6):6719-6748. doi: 10.1021/acs.chemrev.1c00774. Epub 2022 Feb 18. Chem Rev. 2022. PMID: 35179885 Free PMC article. Review.
-
Different states and the associated fates of biomolecular condensates.Essays Biochem. 2022 Dec 16;66(7):849-862. doi: 10.1042/EBC20220054. Essays Biochem. 2022. PMID: 36350032 Free PMC article. Review.
Cited by
-
Phosphatase to kinase switch of a critical enzyme contributes to timing of cell differentiation.mBio. 2024 Jan 16;15(1):e0212523. doi: 10.1128/mbio.02125-23. Epub 2023 Dec 6. mBio. 2024. PMID: 38055339 Free PMC article.
-
Aberrant phase separation is a common killing strategy of positively charged peptides in biology and human disease.bioRxiv [Preprint]. 2023 Mar 9:2023.03.09.531820. doi: 10.1101/2023.03.09.531820. bioRxiv. 2023. PMID: 36945394 Free PMC article. Preprint.
-
Sequence-Dependent Material Properties of Biomolecular Codensates and their Relation to Dilute Phase Conformations.bioRxiv [Preprint]. 2024 Jan 21:2023.05.09.540038. doi: 10.1101/2023.05.09.540038. bioRxiv. 2024. Update in: Nat Commun. 2024 Mar 1;15(1):1912. doi: 10.1038/s41467-024-46223-w. PMID: 37215004 Free PMC article. Updated. Preprint.
-
Protein phase separation: new insights into cell division.Acta Biochim Biophys Sin (Shanghai). 2023 May 30;55(7):1042-1051. doi: 10.3724/abbs.2023093. Acta Biochim Biophys Sin (Shanghai). 2023. PMID: 37249333 Free PMC article.
-
Unlocking the electrochemical functions of biomolecular condensates.Nat Chem Biol. 2024 Nov;20(11):1420-1433. doi: 10.1038/s41589-024-01717-y. Epub 2024 Sep 26. Nat Chem Biol. 2024. PMID: 39327453 Review.
References
-
- Shin, Y. & Brangwynne, C. P. Liquid phase condensation in cell physiology and disease. Science357 (2017). - PubMed
