

The developmentally timed decay of an essential microRNA family is seed-sequence dependent
- PMID: 35947946
- PMCID: PMC9413084
- DOI: 10.1016/j.celrep.2022.111154
The developmentally timed decay of an essential microRNA family is seed-sequence dependent
Abstract
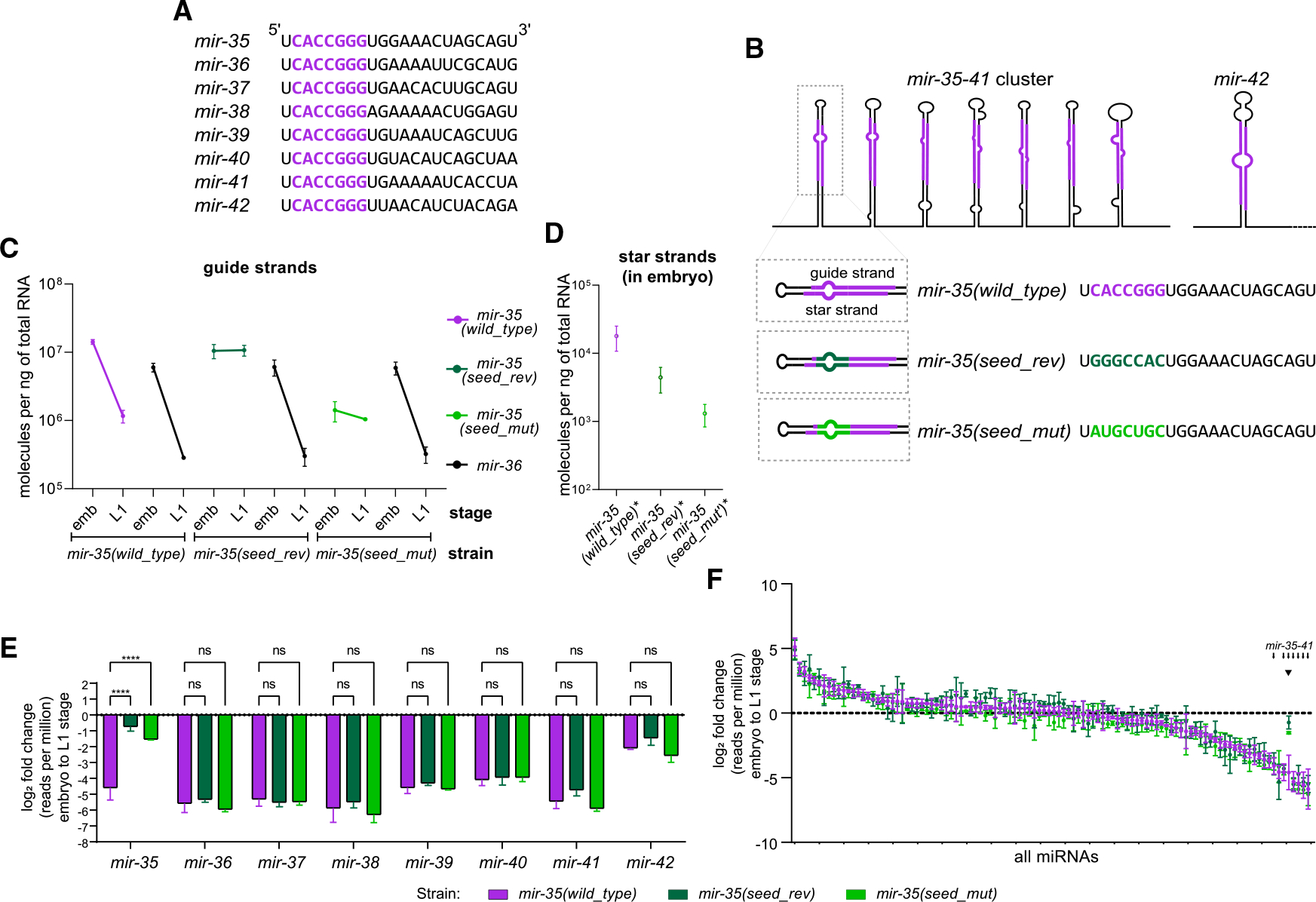
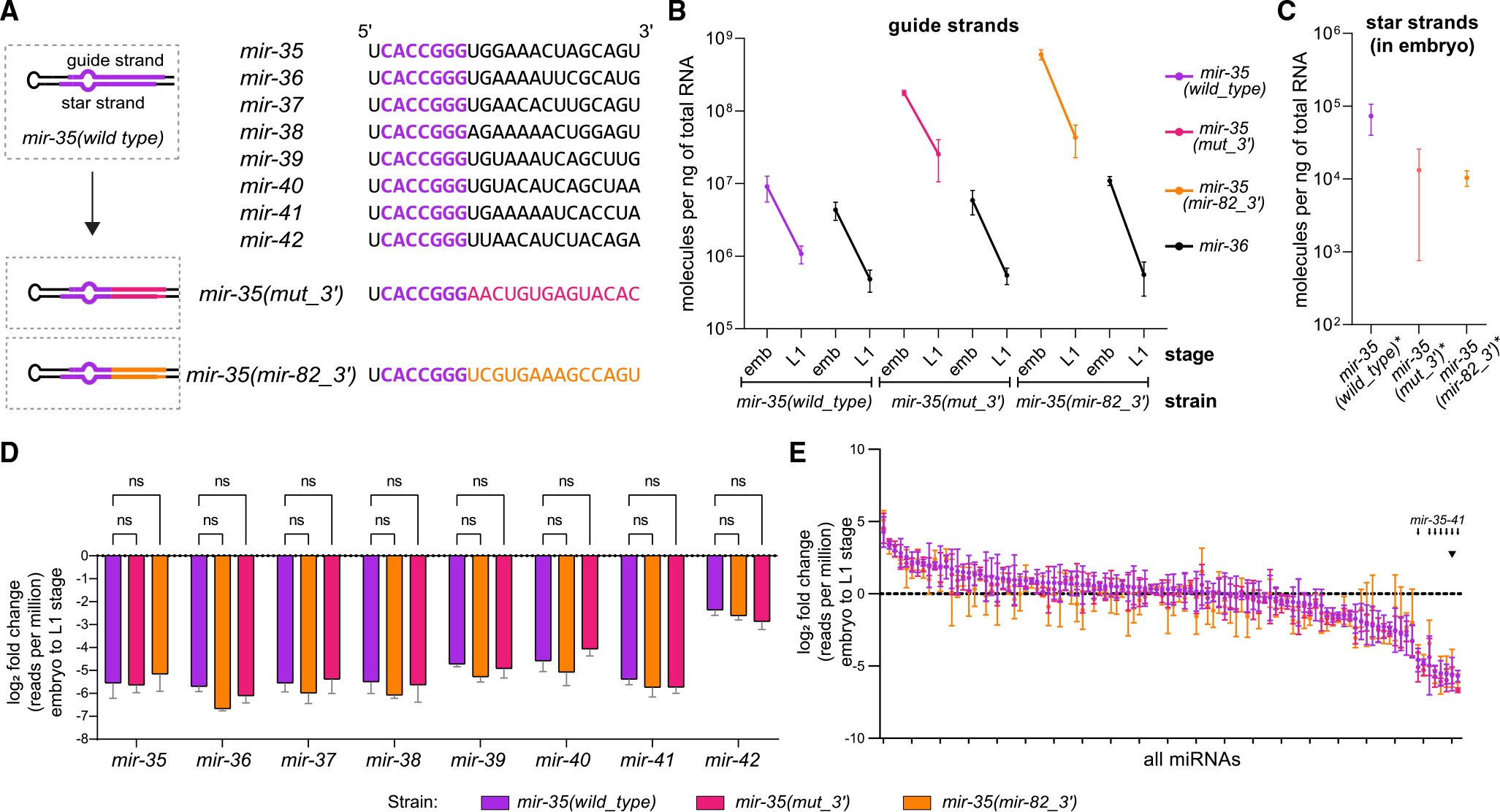
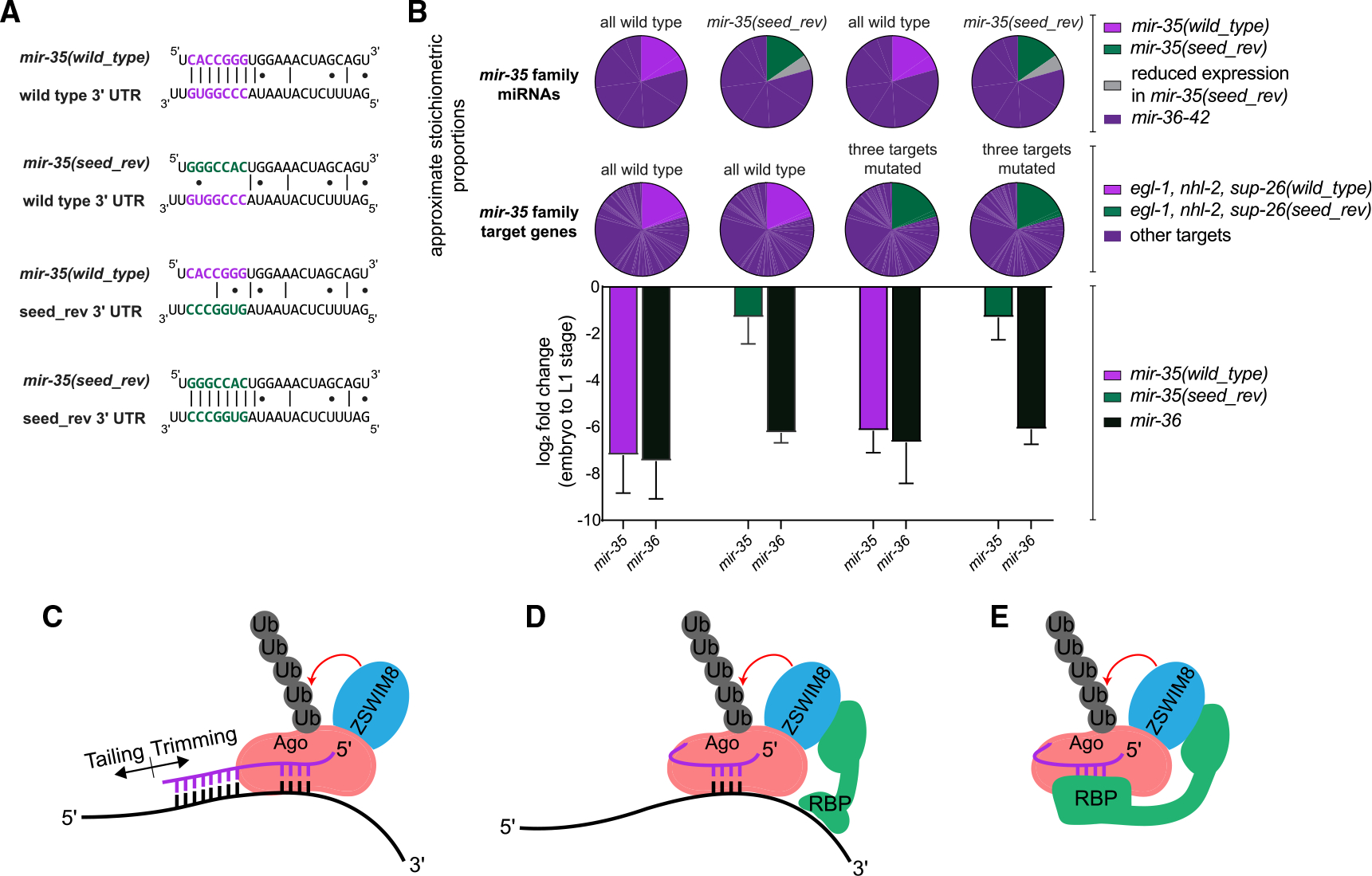
MicroRNA (miRNA) abundance is tightly controlled by regulation of biogenesis and decay. Here, we show that the mir-35 miRNA family undergoes selective decay at the transition from embryonic to larval development in C. elegans. The seed sequence of the miRNA is necessary and largely sufficient for this regulation. Sequences outside the seed (3' end) regulate mir-35 abundance in the embryo but are not necessary for sharp decay at the transition to larval development. Enzymatic modifications of the miRNA 3' end are neither prevalent nor correlated with changes in decay, suggesting that miRNA 3' end display is not a core feature of this mechanism and further supporting a seed-driven decay model. Our findings demonstrate that seed-sequence-specific decay can selectively and coherently regulate all redundant members of a miRNA seed family, a class of mechanism that has great biological and therapeutic potential for dynamic regulation of a miRNA family's target repertoire.
Keywords: C. elegans; CP: Cell biology; CP: Developmental biology; EBAX-1; ZSWIM8; developmental transitions; embryonic development; miRNA; miRNA decay; miRNA degradation; miRNA tailing; miRNA turnover; microRNA; mir-35; mir-35 family; mir-35-41; post-transcriptional regulation; target-directed miRNA degradation; TDMD.
Published by Elsevier Inc.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures

Similar articles
-
Cooperative target mRNA destabilization and translation inhibition by miR-58 microRNA family in C. elegans.Genome Res. 2015 Nov;25(11):1680-91. doi: 10.1101/gr.183160.114. Epub 2015 Jul 31. Genome Res. 2015. PMID: 26232411 Free PMC article.
-
Screening by deep sequencing reveals mediators of microRNA tailing in C. elegans.Nucleic Acids Res. 2021 Nov 8;49(19):11167-11180. doi: 10.1093/nar/gkab840. Nucleic Acids Res. 2021. PMID: 34586415 Free PMC article.
-
Active turnover modulates mature microRNA activity in Caenorhabditis elegans.Nature. 2009 Sep 24;461(7263):546-9. doi: 10.1038/nature08349. Epub 2009 Sep 6. Nature. 2009. PMID: 19734881
-
Target-directed microRNA degradation: Mechanisms, significance, and functional implications.Wiley Interdiscip Rev RNA. 2024 Mar-Apr;15(2):e1832. doi: 10.1002/wrna.1832. Wiley Interdiscip Rev RNA. 2024. PMID: 38448799 Free PMC article. Review.
-
Maternal effects of microRNAs in early embryogenesis.RNA Biol. 2018 Feb 1;15(2):165-169. doi: 10.1080/15476286.2017.1402999. Epub 2017 Dec 8. RNA Biol. 2018. PMID: 29120257 Free PMC article. Review.
Cited by
-
MicroRNA turnover: a tale of tailing, trimming, and targets.Trends Biochem Sci. 2023 Jan;48(1):26-39. doi: 10.1016/j.tibs.2022.06.005. Epub 2022 Jul 7. Trends Biochem Sci. 2023. PMID: 35811249 Free PMC article. Review.
-
Screening of Drosophila microRNA-degradation sequences reveals Argonaute1 mRNA's role in regulating miR-999.Nat Commun. 2023 Apr 13;14(1):2108. doi: 10.1038/s41467-023-37819-9. Nat Commun. 2023. PMID: 37055443 Free PMC article.
-
To kill a microRNA: emerging concepts in target-directed microRNA degradation.Nucleic Acids Res. 2024 Feb 28;52(4):1558-1574. doi: 10.1093/nar/gkae003. Nucleic Acids Res. 2024. PMID: 38224449 Free PMC article. Review.
-
Pseudogene GSTM3P1 derived long non-coding RNA promotes ischemic acute kidney injury by target directed microRNA degradation of kidney-protective mir-668.Kidney Int. 2024 Oct;106(4):640-657. doi: 10.1016/j.kint.2024.06.027. Epub 2024 Jul 27. Kidney Int. 2024. PMID: 39074555
-
The miRNA-target interactions: An underestimated intricacy.Nucleic Acids Res. 2024 Feb 28;52(4):1544-1557. doi: 10.1093/nar/gkad1142. Nucleic Acids Res. 2024. PMID: 38033323 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
