

Critical contribution of 3' non-seed base pairing to the in vivo function of the evolutionarily conserved let-7a microRNA
- PMID: 35476978
- PMCID: PMC9161110
- DOI: 10.1016/j.celrep.2022.110745
Critical contribution of 3' non-seed base pairing to the in vivo function of the evolutionarily conserved let-7a microRNA
Abstract
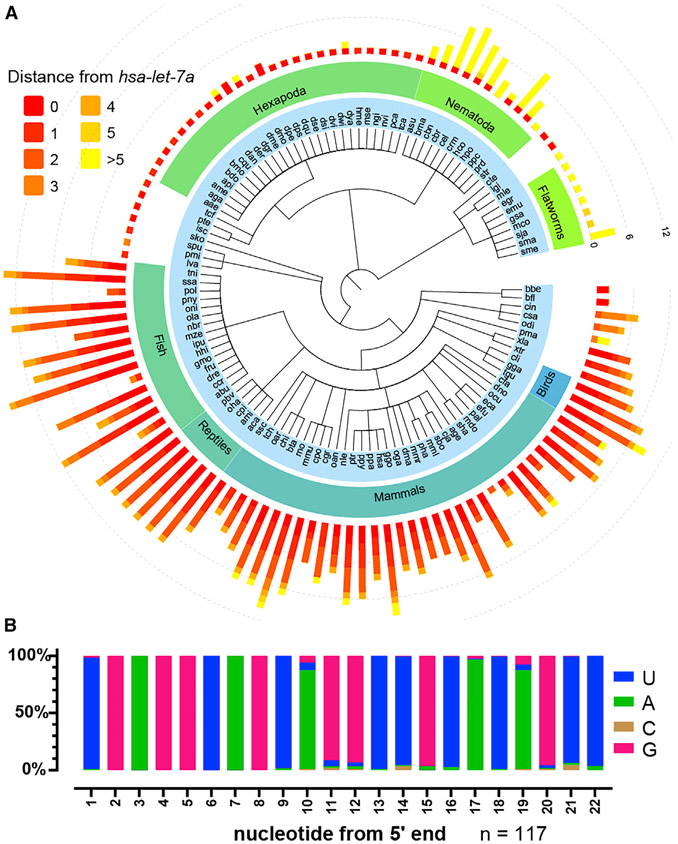
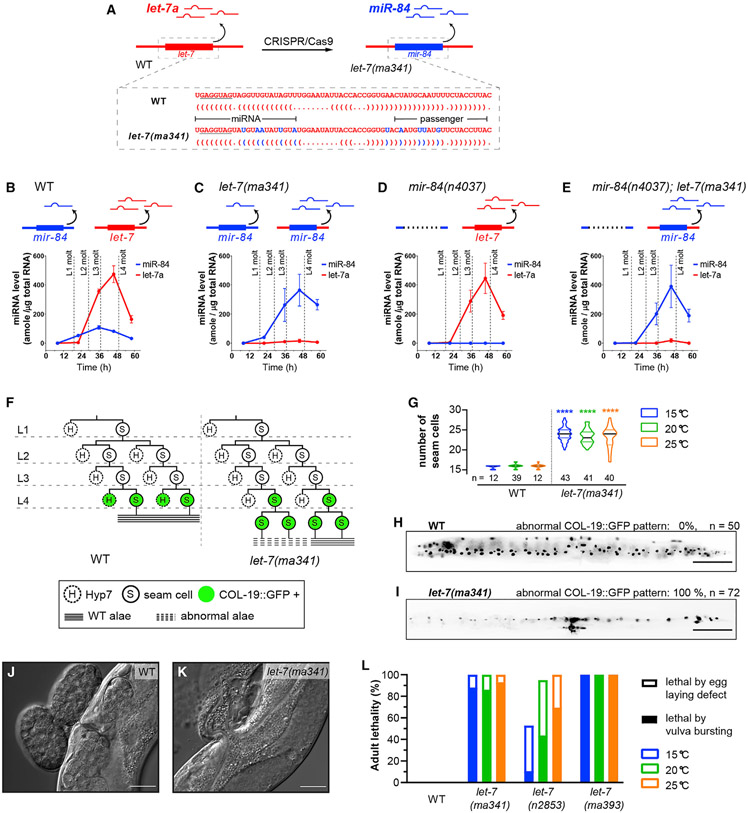
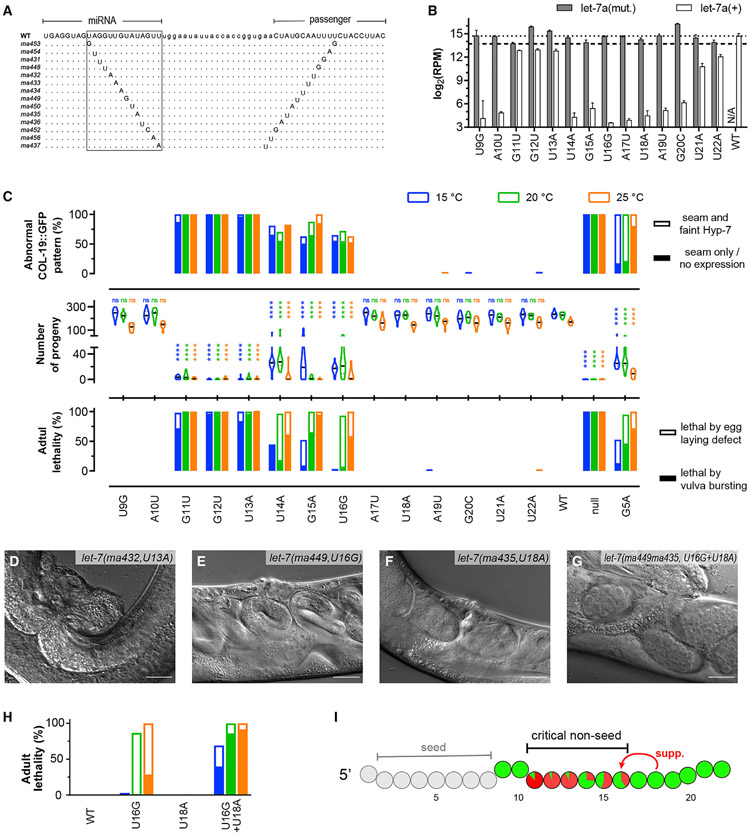
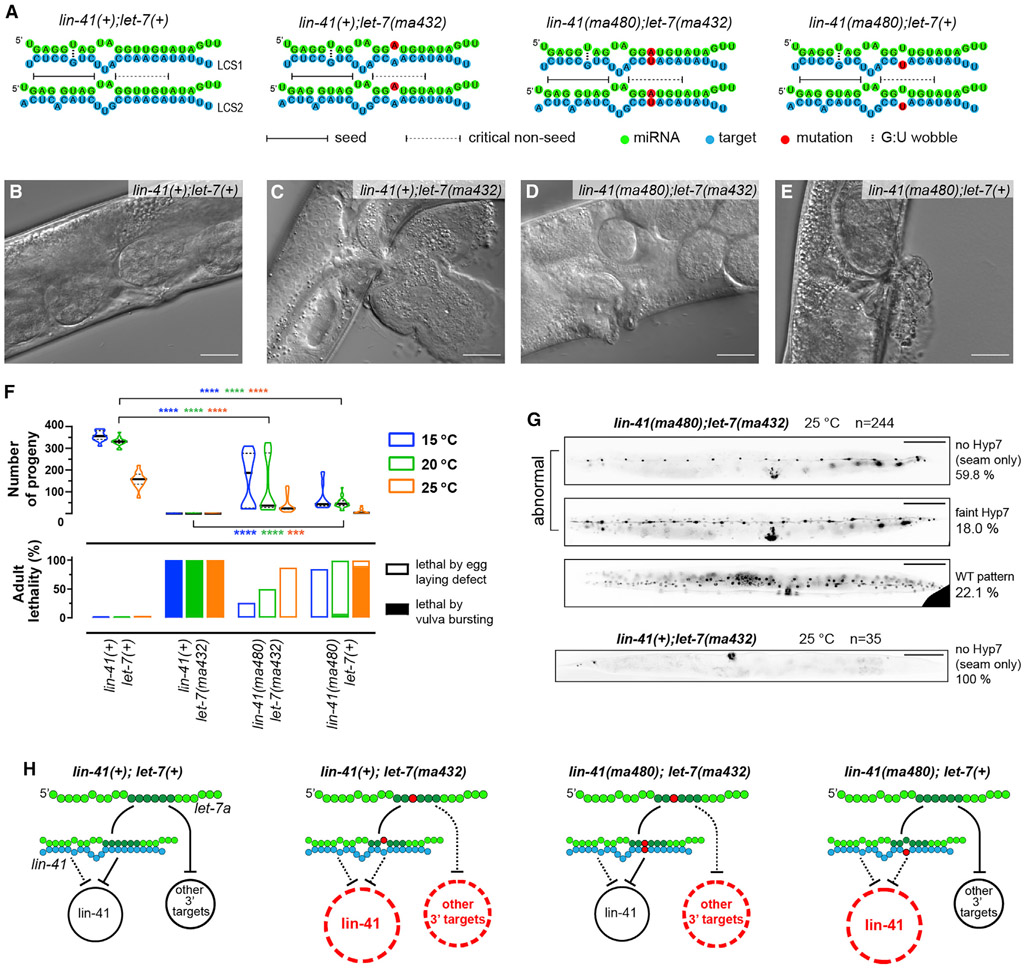
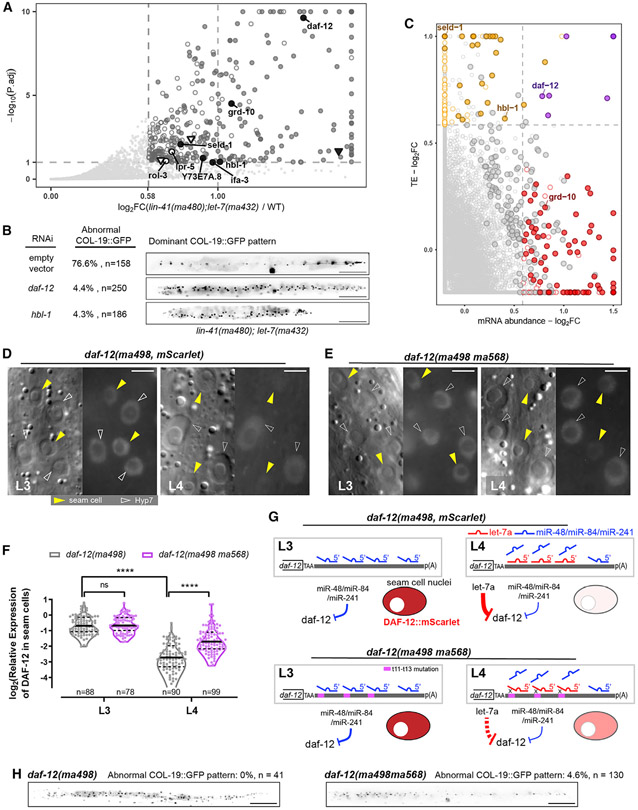
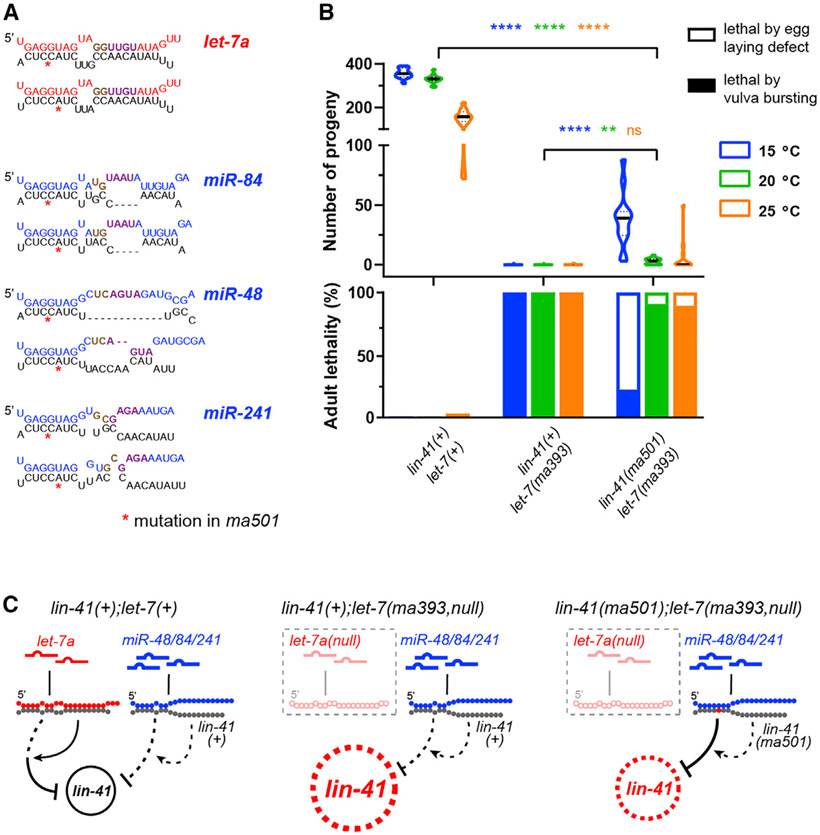
Base pairing of the seed region (g2-g8) is essential for microRNA targeting; however, the in vivo function of the 3' non-seed region (g9-g22) is less well understood. Here, we report a systematic investigation of the in vivo roles of 3' non-seed nucleotides in microRNA let-7a, whose entire g9-g22 region is conserved among bilaterians. We find that the 3' non-seed sequence functionally distinguishes let-7a from its family paralogs. The complete pairing of g11-g16 is essential for let-7a to fully repress multiple key targets, including evolutionarily conserved lin-41, daf-12, and hbl-1. Nucleotides at g17-g22 are less critical but may compensate for mismatches in the g11-g16 region. Interestingly, a certain minimal complementarity to let-7a 3' non-seed sequence can be required even for sites with perfect seed pairing. These results provide evidence that the specific configurations of both seed and 3' non-seed base pairing can critically influence microRNA-mediated gene regulation in vivo.
Keywords: 3′ non-seed pairing; CP: Molecular biology; daf-12; let-7; lin-41; microRNA; post-transcriptional regulation.
Copyright © 2022 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures

Similar articles
-
Efficient 3'-pairing renders microRNA targeting less sensitive to mRNA seed accessibility.Nucleic Acids Res. 2023 Nov 10;51(20):11162-11177. doi: 10.1093/nar/gkad795. Nucleic Acids Res. 2023. PMID: 37819016 Free PMC article.
-
Complementarity to an miRNA seed region is sufficient to induce moderate repression of a target transcript in the unicellular green alga Chlamydomonas reinhardtii.Plant J. 2013 Dec;76(6):1045-56. doi: 10.1111/tpj.12354. Plant J. 2013. PMID: 24127635
-
miRNA Targeting: Growing beyond the Seed.Trends Genet. 2019 Mar;35(3):215-222. doi: 10.1016/j.tig.2018.12.005. Epub 2019 Jan 9. Trends Genet. 2019. PMID: 30638669 Free PMC article. Review.
-
Expanding the microRNA targeting code: functional sites with centered pairing.Mol Cell. 2010 Jun 25;38(6):789-802. doi: 10.1016/j.molcel.2010.06.005. Mol Cell. 2010. PMID: 20620952 Free PMC article.
-
The role of Myc and let-7a in glioblastoma, glucose metabolism and response to therapy.Arch Biochem Biophys. 2015 Aug 15;580:84-92. doi: 10.1016/j.abb.2015.07.005. Epub 2015 Jul 4. Arch Biochem Biophys. 2015. PMID: 26151775 Review.
Cited by
-
To kill a microRNA: emerging concepts in target-directed microRNA degradation.Nucleic Acids Res. 2024 Feb 28;52(4):1558-1574. doi: 10.1093/nar/gkae003. Nucleic Acids Res. 2024. PMID: 38224449 Free PMC article. Review.
-
Transcriptome-Wide Analysis of microRNA-mRNA Correlations in Tissue Identifies microRNA Targeting Determinants.Noncoding RNA. 2023 Feb 13;9(1):15. doi: 10.3390/ncrna9010015. Noncoding RNA. 2023. PMID: 36827548 Free PMC article.
-
Recent advances in understanding microRNA function and regulation in C. elegans.Semin Cell Dev Biol. 2024 Feb 15;154(Pt A):4-13. doi: 10.1016/j.semcdb.2023.03.011. Epub 2023 Apr 11. Semin Cell Dev Biol. 2024. PMID: 37055330 Review.
-
Efficient 3'-pairing renders microRNA targeting less sensitive to mRNA seed accessibility.Nucleic Acids Res. 2023 Nov 10;51(20):11162-11177. doi: 10.1093/nar/gkad795. Nucleic Acids Res. 2023. PMID: 37819016 Free PMC article.
-
Expression, not sequence, distinguishes miR-238 from its miR-239ab sister miRNAs in promoting longevity in Caenorhabditis elegans.PLoS Genet. 2023 Nov 27;19(11):e1011055. doi: 10.1371/journal.pgen.1011055. eCollection 2023 Nov. PLoS Genet. 2023. PMID: 38011256 Free PMC article.
References
-
- Abrahante JE, Daul AL, Li M, Volk ML, Tennessen JM, Miller EA, and Rougvie AE (2003). The Caenorhabditis elegans hunchback-like gene lin-57/hbl-1 controls developmental time and is regulated by microRNAs. Dev. Cell 4, 625–637. - PubMed
-
- Aeschimann F, Kumari P, Bartake H, Gaidatzis D, Xu L, Ciosk R, and Grosshans H (2017). LIN41 post-transcriptionally silences mRNAs by two distinct and position-dependent mechanisms. Mol. Cell 65, 476–489.e4. - PubMed
-
- Aeschimann F, Xiong J, Arnold A, Dieterich C, and Grosshans H (2015). Transcriptome-wide measurement of ribosomal occupancy by ribosome profiling. Methods 85, 75–89. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
