

On the role of phase separation in the biogenesis of membraneless compartments
- PMID: 35107832
- PMCID: PMC8886532
- DOI: 10.15252/embj.2021109952
On the role of phase separation in the biogenesis of membraneless compartments
Abstract
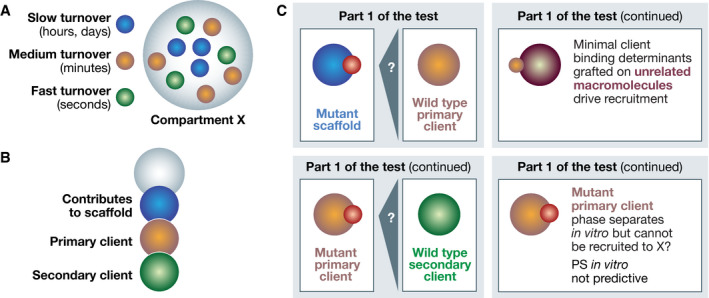
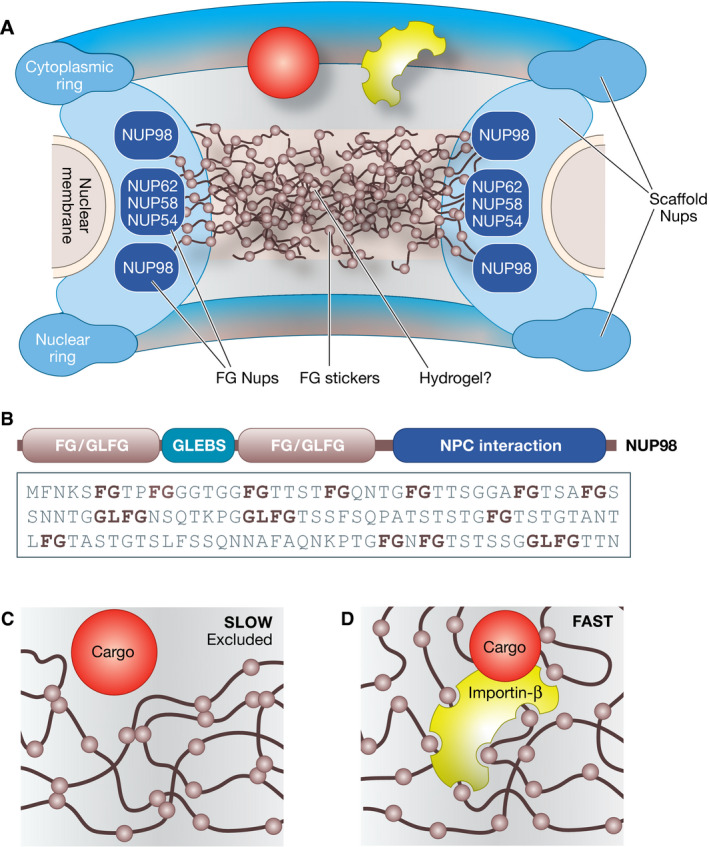
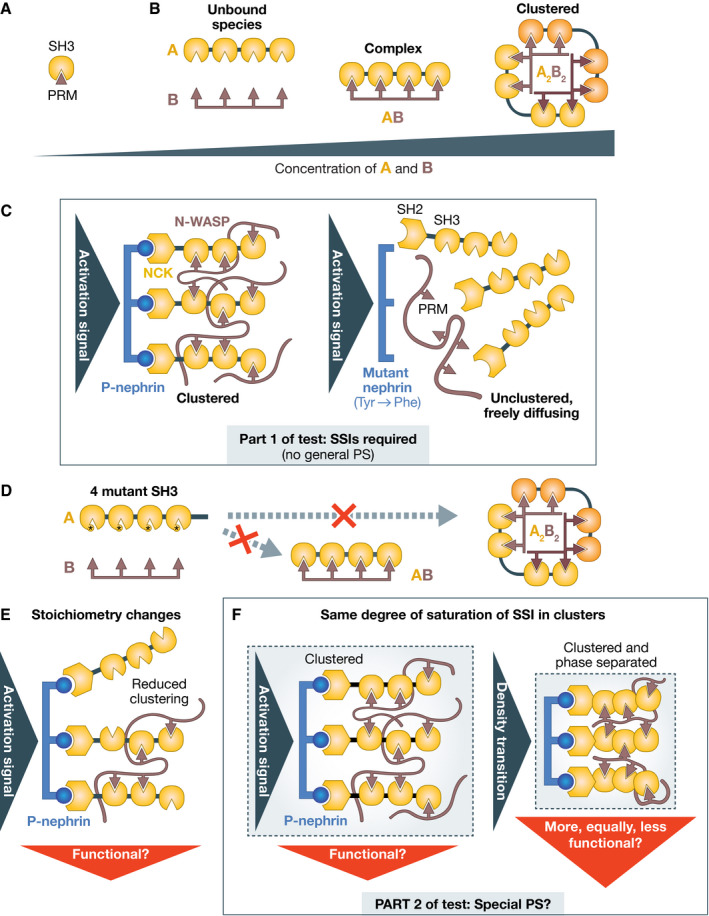
Molecular mechanistic biology has ushered us into the world of life's building blocks, revealing their interactions in macromolecular complexes and inspiring strategies for detailed functional interrogations. The biogenesis of membraneless cellular compartments, functional mesoscale subcellular locales devoid of strong internal order and delimiting membranes, is among mechanistic biology's most demanding current challenges. A developing paradigm, biomolecular phase separation, emphasizes solvation of the building blocks through low-affinity, weakly adhesive unspecific interactions as the driver of biogenesis of membraneless compartments. Here, I discuss the molecular underpinnings of the phase separation paradigm and demonstrate that validating its assumptions is much more challenging than hitherto appreciated. I also discuss that highly specific interactions, rather than unspecific ones, appear to be the main driver of biogenesis of subcellular compartments, while phase separation may be harnessed locally in selected instances to generate material properties tailored for specific functions, as exemplified by nucleocytoplasmic transport.
© 2022 The Author. Published under the terms of the CC BY 4.0 license.
Figures
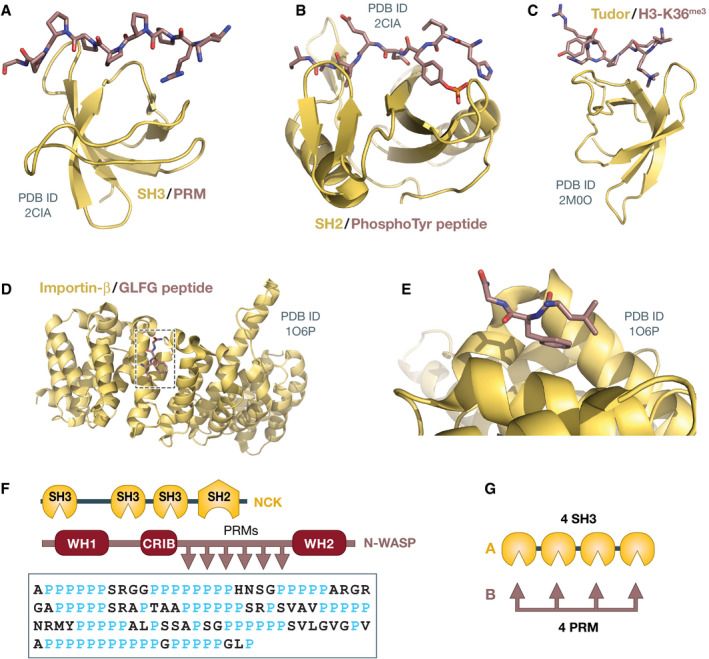
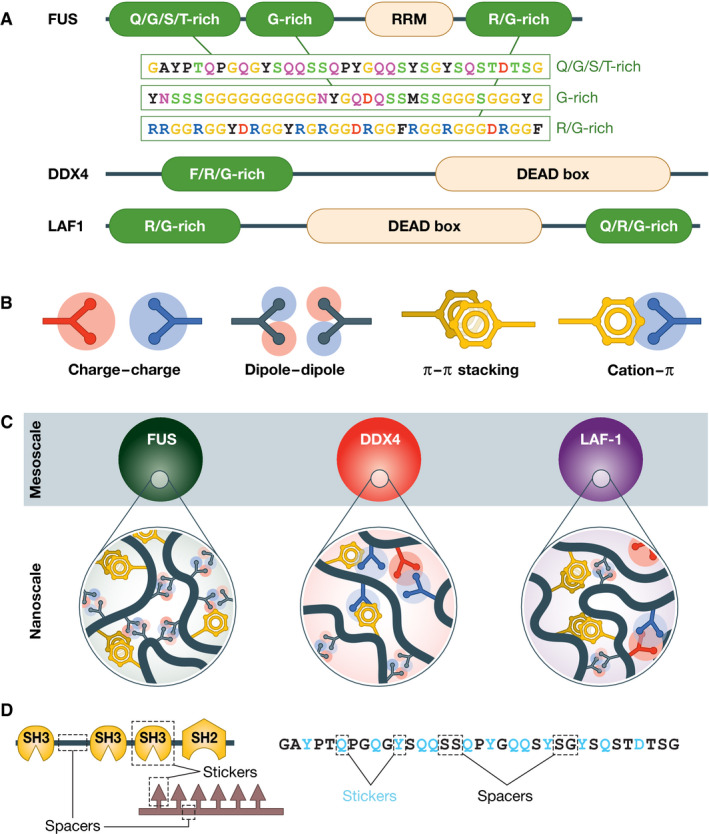
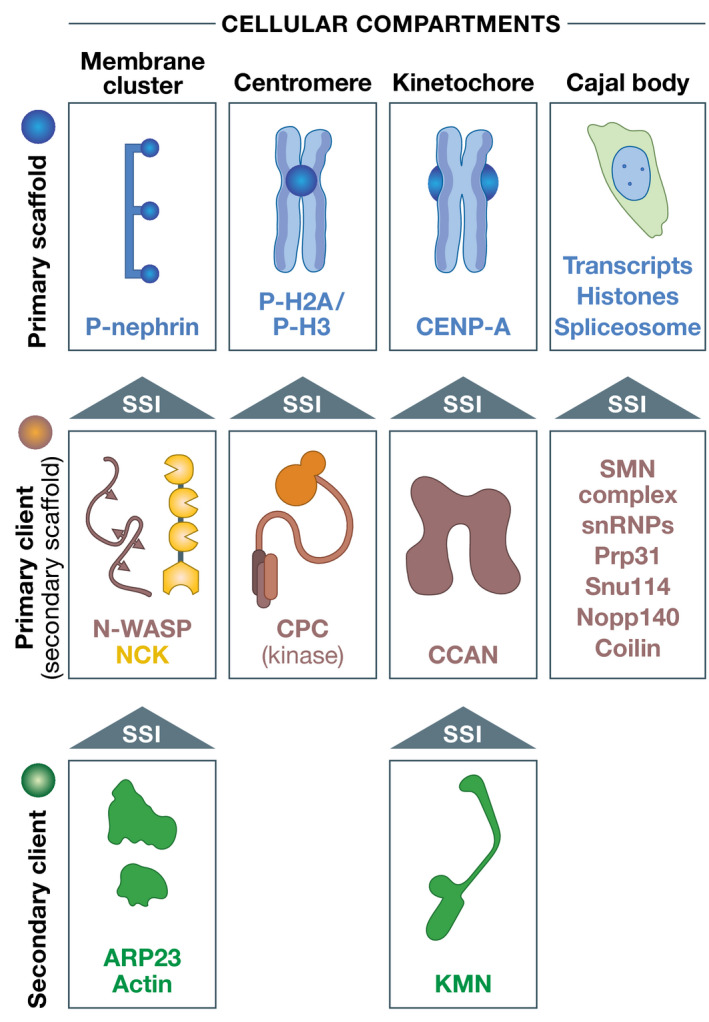
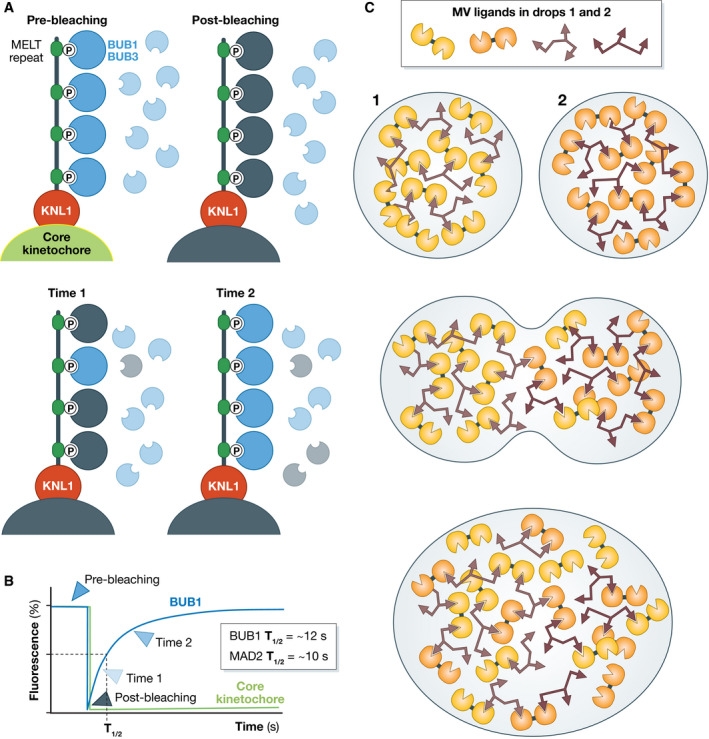
Similar articles
-
Phase Separation in Membrane Biology: The Interplay between Membrane-Bound Organelles and Membraneless Condensates.Dev Cell. 2020 Oct 12;55(1):30-44. doi: 10.1016/j.devcel.2020.06.033. Epub 2020 Jul 28. Dev Cell. 2020. PMID: 32726575 Review.
-
Hyperosmotic phase separation: Condensates beyond inclusions, granules and organelles.J Biol Chem. 2021 Jan-Jun;296:100044. doi: 10.1074/jbc.REV120.010899. Epub 2020 Nov 23. J Biol Chem. 2021. PMID: 33168632 Free PMC article. Review.
-
Membraneless nuclear organelles and the search for phases within phases.Wiley Interdiscip Rev RNA. 2019 Mar;10(2):e1514. doi: 10.1002/wrna.1514. Epub 2018 Oct 25. Wiley Interdiscip Rev RNA. 2019. PMID: 30362243 Review.
-
Three archetypical classes of macromolecular regulators of protein liquid-liquid phase separation.Proc Natl Acad Sci U S A. 2019 Sep 24;116(39):19474-19483. doi: 10.1073/pnas.1907849116. Epub 2019 Sep 10. Proc Natl Acad Sci U S A. 2019. PMID: 31506351 Free PMC article.
-
Phase transition of a disordered nuage protein generates environmentally responsive membraneless organelles.Mol Cell. 2015 Mar 5;57(5):936-947. doi: 10.1016/j.molcel.2015.01.013. Mol Cell. 2015. PMID: 25747659 Free PMC article.
Cited by
-
Malaria parasite centrins can assemble by Ca2+-inducible condensation.PLoS Pathog. 2023 Dec 27;19(12):e1011899. doi: 10.1371/journal.ppat.1011899. eCollection 2023 Dec. PLoS Pathog. 2023. PMID: 38150475 Free PMC article.
-
Illuminating Intrinsically Disordered Proteins with Integrative Structural Biology.Biomolecules. 2023 Jan 7;13(1):124. doi: 10.3390/biom13010124. Biomolecules. 2023. PMID: 36671509 Free PMC article. Review.
-
The relative binding position of Nck and Grb2 adaptors impacts actin-based motility of Vaccinia virus.Elife. 2022 Jul 7;11:e74655. doi: 10.7554/eLife.74655. Elife. 2022. PMID: 35796545 Free PMC article.
-
Building the synaptonemal complex: Molecular interactions between the axis and the central region.PLoS Genet. 2023 Jul 20;19(7):e1010822. doi: 10.1371/journal.pgen.1010822. eCollection 2023 Jul. PLoS Genet. 2023. PMID: 37471284 Free PMC article. Review.
-
Biological condensates form percolated networks with molecular motion properties distinctly different from dilute solutions.Elife. 2023 Jun 1;12:e81907. doi: 10.7554/eLife.81907. Elife. 2023. PMID: 37261897 Free PMC article.
References
-
- Alberti S, Hyman AA (2021) Biomolecular condensates at the nexus of cellular stress, protein aggregation disease and ageing. Nat Rev Mol Cell Biol 22: 196–213 - PubMed
-
- Aloy P, Ceulemans H, Stark A, Russell RB (2003) The relationship between sequence and interaction divergence in proteins. J Mol Biol 332: 989–998 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
