

Nuclear dengue virus NS5 antagonizes expression of PAF1-dependent immune response genes
- PMID: 34797876
- PMCID: PMC8641875
- DOI: 10.1371/journal.ppat.1010100
Nuclear dengue virus NS5 antagonizes expression of PAF1-dependent immune response genes
Abstract
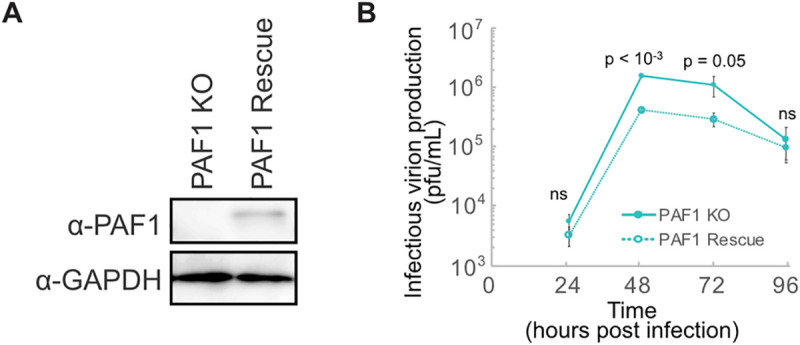
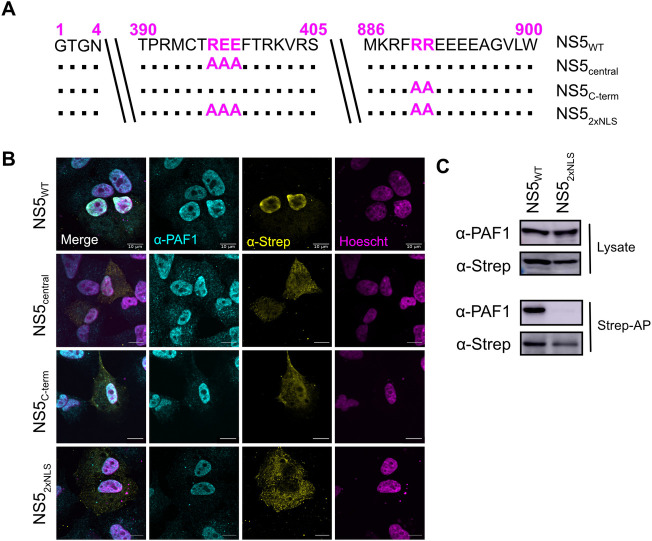
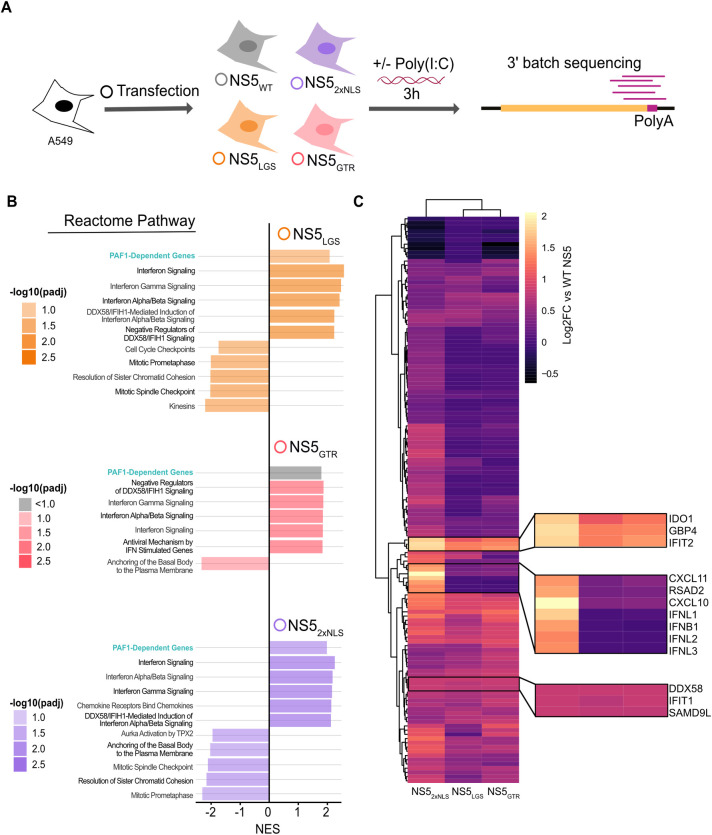
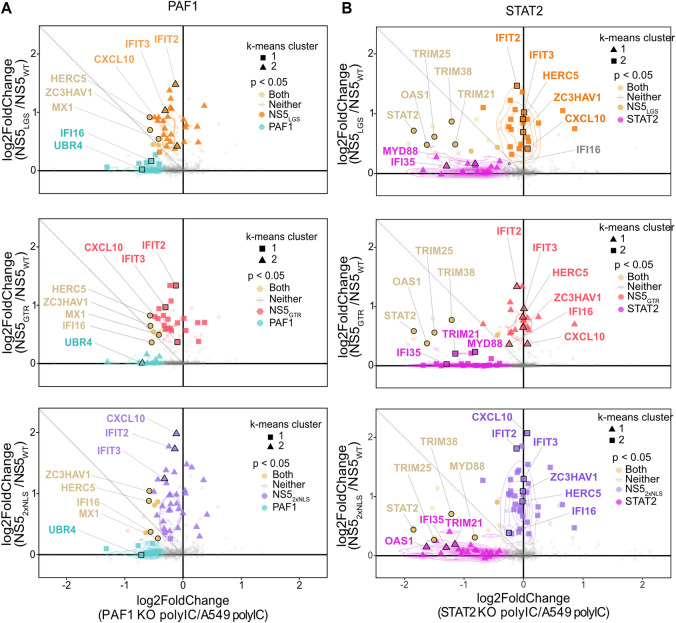
Dengue virus (DENV) disruption of the innate immune response is critical to establish infection. DENV non-structural protein 5 (NS5) plays a central role in this disruption, such as antagonism of STAT2. We recently found that DENV serotype 2 (DENV2) NS5 interacts with Polymerase associated factor 1 complex (PAF1C). The primary members of PAF1C are PAF1, LEO1, CTR9, and CDC73. This nuclear complex is an emerging player in the immune response. It promotes the expression of many genes, including genes related to the antiviral, antimicrobial and inflammatory responses, through close association with the chromatin of these genes. Our previous work demonstrated that NS5 antagonizes PAF1C recruitment to immune response genes. However, it remains unknown if NS5 antagonism of PAF1C is complementary to its antagonism of STAT2. Here, we show that knockout of PAF1 enhances DENV2 infectious virion production. By comparing gene expression profiles in PAF1 and STAT2 knockout cells, we find that PAF1 is necessary to express immune response genes that are STAT2-independent. Finally, we mapped the viral determinants for the NS5-PAF1C protein interaction. We found that NS5 nuclear localization and the C-terminal region of the methyltransferase domain are required for its interaction with PAF1C. Mutation of these regions rescued the expression of PAF1-dependent immune response genes that are antagonized by NS5. In sum, our results support a role for PAF1C in restricting DENV2 replication that NS5 antagonizes through its protein interaction with PAF1C.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures



Similar articles
-
SUMO Modification Stabilizes Dengue Virus Nonstructural Protein 5 To Support Virus Replication.J Virol. 2016 Apr 14;90(9):4308-4319. doi: 10.1128/JVI.00223-16. Print 2016 May. J Virol. 2016. PMID: 26889037 Free PMC article.
-
A cellular screening platform, stably expressing DENV2 NS5, defines a novel anti-DENV mechanism of action of Apigenin based on STAT2 activation.Virology. 2023 Jun;583:1-13. doi: 10.1016/j.virol.2023.03.016. Epub 2023 Apr 5. Virology. 2023. PMID: 37060797
-
NS5 Sumoylation Directs Nuclear Responses That Permit Zika Virus To Persistently Infect Human Brain Microvascular Endothelial Cells.J Virol. 2020 Sep 15;94(19):e01086-20. doi: 10.1128/JVI.01086-20. Print 2020 Sep 15. J Virol. 2020. PMID: 32699085 Free PMC article.
-
The Transactions of NS3 and NS5 in Flaviviral RNA Replication.Adv Exp Med Biol. 2018;1062:147-163. doi: 10.1007/978-981-10-8727-1_11. Adv Exp Med Biol. 2018. PMID: 29845531 Review.
-
Innate immunity evasion by Dengue virus.Viruses. 2012 Mar;4(3):397-413. doi: 10.3390/v4030397. Epub 2012 Mar 15. Viruses. 2012. PMID: 22590678 Free PMC article. Review.
Cited by
-
TMEM41B and VMP1 modulate cellular lipid and energy metabolism for facilitating dengue virus infection.PLoS Pathog. 2022 Aug 8;18(8):e1010763. doi: 10.1371/journal.ppat.1010763. eCollection 2022 Aug. PLoS Pathog. 2022. PMID: 35939522 Free PMC article.
-
Genomic Characterization of Dengue Virus Outbreak in 2022 from Pakistan.Vaccines (Basel). 2023 Jan 11;11(1):163. doi: 10.3390/vaccines11010163. Vaccines (Basel). 2023. PMID: 36680008 Free PMC article.
-
The innate immune factor RPRD2/REAF and its role in the Lv2 restriction of HIV.mBio. 2023 Dec 19;14(6):e0257221. doi: 10.1128/mbio.02572-21. Epub 2023 Oct 26. mBio. 2023. PMID: 37882563 Free PMC article. Review.
-
A Zika virus protein expression screen in Drosophila to investigate targeted host pathways during development.Dis Model Mech. 2024 Feb 1;17(2):dmm050297. doi: 10.1242/dmm.050297. Epub 2024 Feb 28. Dis Model Mech. 2024. PMID: 38214058 Free PMC article.
-
Interaction of HDAC2 with SARS-CoV-2 NSP5 and IRF3 Is Not Required for NSP5-Mediated Inhibition of Type I Interferon Signaling Pathway.Microbiol Spectr. 2022 Oct 26;10(5):e0232222. doi: 10.1128/spectrum.02322-22. Epub 2022 Sep 29. Microbiol Spectr. 2022. PMID: 36173315 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
