

PDZD-8 and TEX-2 regulate endosomal PI(4,5)P2 homeostasis via lipid transport to promote embryogenesis in C. elegans
- PMID: 34663803
- PMCID: PMC8523718
- DOI: 10.1038/s41467-021-26177-z
PDZD-8 and TEX-2 regulate endosomal PI(4,5)P2 homeostasis via lipid transport to promote embryogenesis in C. elegans
Abstract
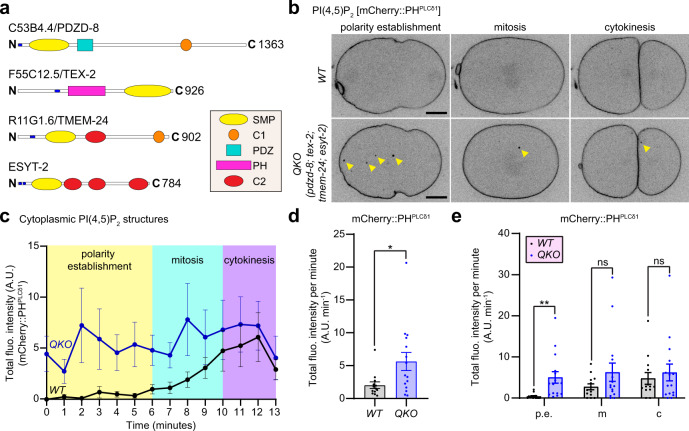
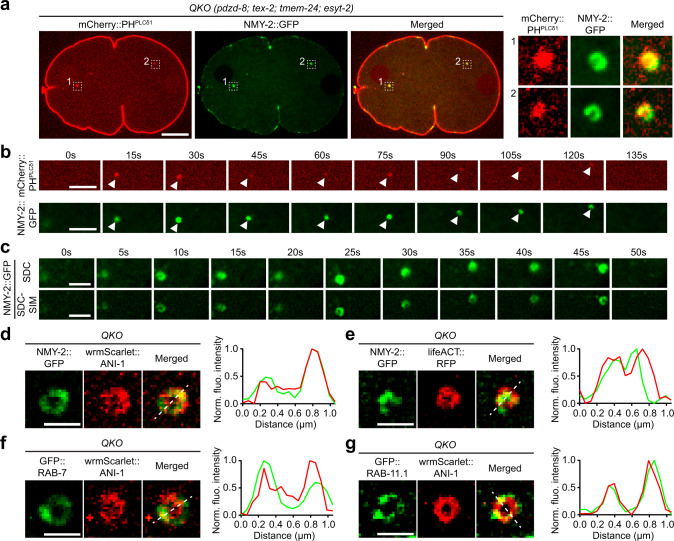
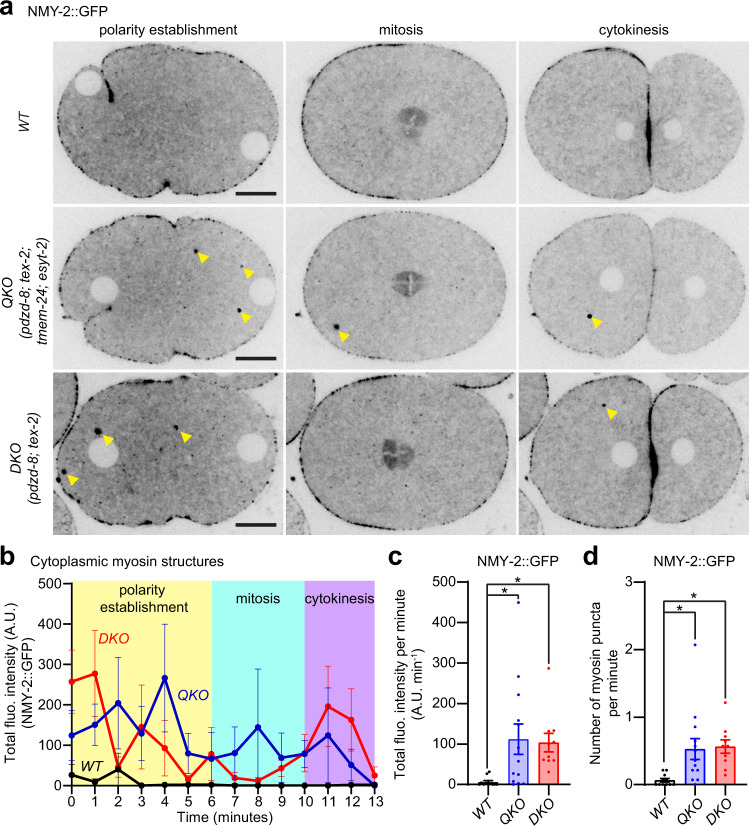
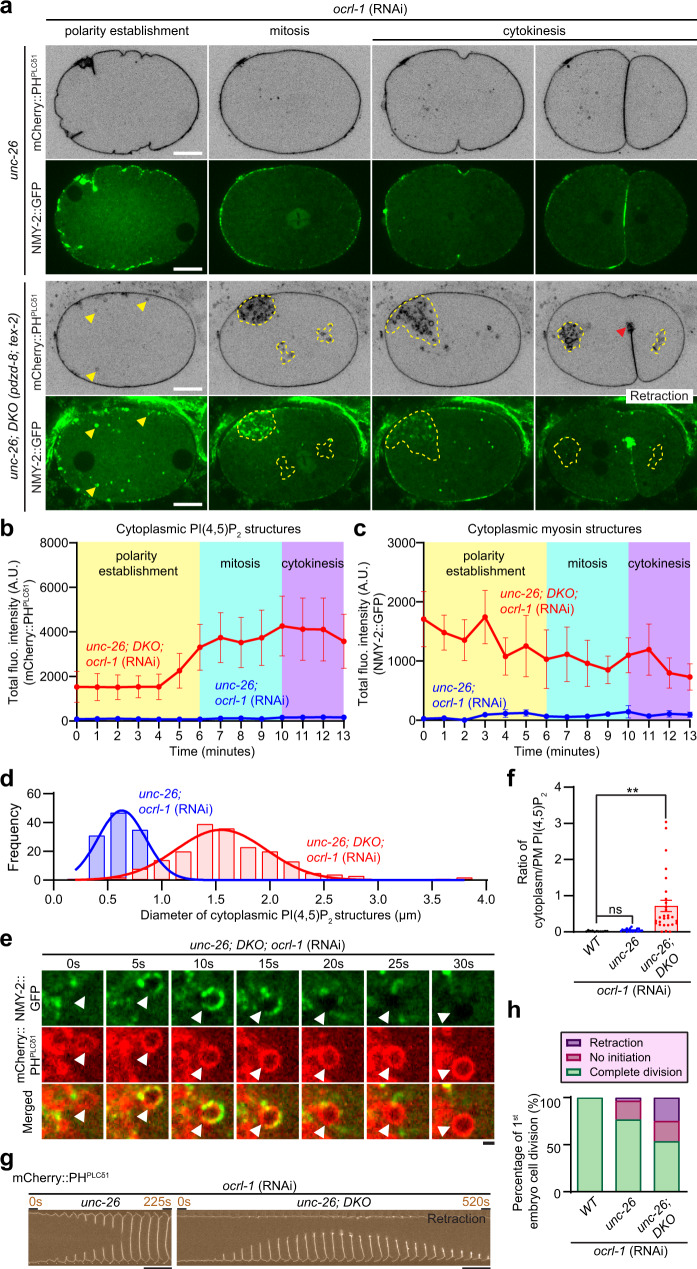
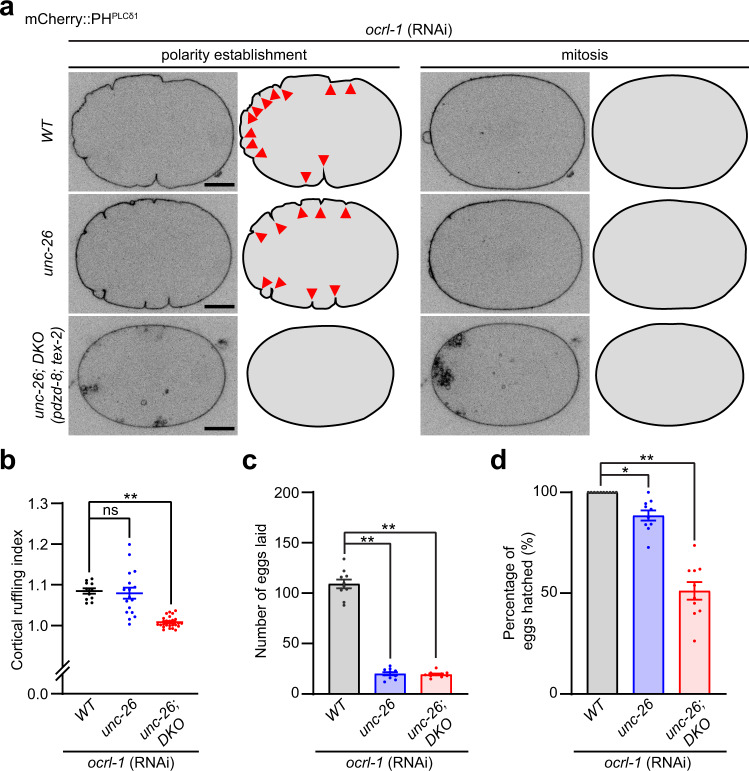
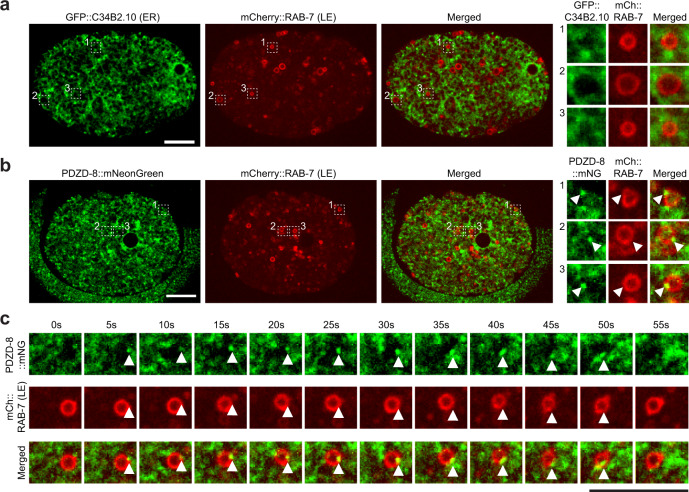
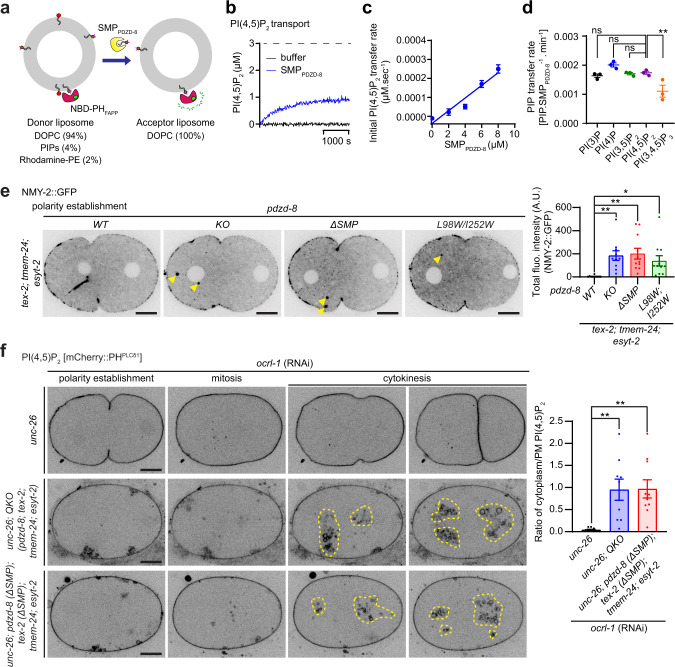
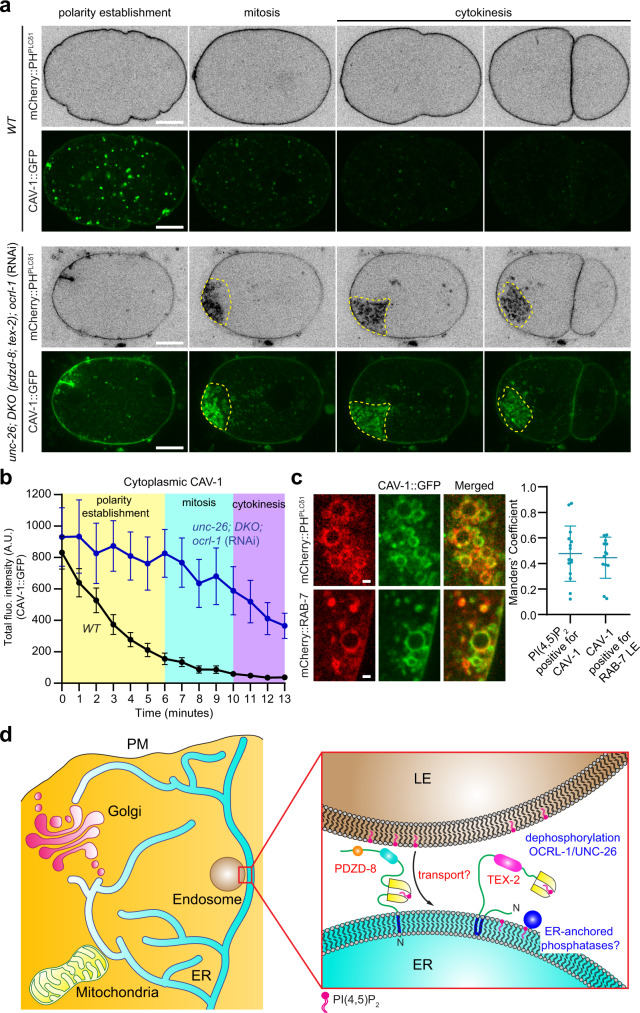
Different types of cellular membranes have unique lipid compositions that are important for their functional identity. PI(4,5)P2 is enriched in the plasma membrane where it contributes to local activation of key cellular events, including actomyosin contraction and cytokinesis. However, how cells prevent PI(4,5)P2 from accumulating in intracellular membrane compartments, despite constant intermixing and exchange of lipid membranes, is poorly understood. Using the C. elegans early embryo as our model system, we show that the evolutionarily conserved lipid transfer proteins, PDZD-8 and TEX-2, act together with the PI(4,5)P2 phosphatases, OCRL-1 and UNC-26/synaptojanin, to prevent the build-up of PI(4,5)P2 on endosomal membranes. In the absence of these four proteins, large amounts of PI(4,5)P2 accumulate on endosomes, leading to embryonic lethality due to ectopic recruitment of proteins involved in actomyosin contractility. PDZD-8 localizes to the endoplasmic reticulum and regulates endosomal PI(4,5)P2 levels via its lipid harboring SMP domain. Accumulation of PI(4,5)P2 on endosomes is accompanied by impairment of their degradative capacity. Thus, cells use multiple redundant systems to maintain endosomal PI(4,5)P2 homeostasis.
© 2021. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
RAB-10-GTPase-mediated regulation of endosomal phosphatidylinositol-4,5-bisphosphate.Proc Natl Acad Sci U S A. 2012 Aug 28;109(35):E2306-15. doi: 10.1073/pnas.1205278109. Epub 2012 Aug 6. Proc Natl Acad Sci U S A. 2012. PMID: 22869721 Free PMC article.
-
RdgBα reciprocally transfers PA and PI at ER-PM contact sites to maintain PI(4,5)P2 homoeostasis during phospholipase C signalling in Drosophila photoreceptors.Biochem Soc Trans. 2016 Feb;44(1):286-92. doi: 10.1042/BST20150228. Biochem Soc Trans. 2016. PMID: 26862217 Review.
-
RACK-1 directs dynactin-dependent RAB-11 endosomal recycling during mitosis in Caenorhabditis elegans.Mol Biol Cell. 2009 Mar;20(6):1629-38. doi: 10.1091/mbc.e08-09-0917. Epub 2009 Jan 21. Mol Biol Cell. 2009. PMID: 19158384 Free PMC article.
-
Interactions between Rab and Arf GTPases regulate endosomal phosphatidylinositol-4,5-bisphosphate during endocytic recycling.Small GTPases. 2013 Apr-Jun;4(2):106-9. doi: 10.4161/sgtp.23477. Epub 2013 Feb 7. Small GTPases. 2013. PMID: 23392104 Free PMC article.
-
Cholesterol transfer at endosomal-organelle membrane contact sites.Curr Opin Lipidol. 2018 Jun;29(3):212-217. doi: 10.1097/MOL.0000000000000506. Curr Opin Lipidol. 2018. PMID: 29629999 Review.
Cited by
-
Recent Advances on Synaptotagmin-Like Mitochondrial-Lipid Binding Protein Domain Containing Lipid Transfer Proteins.Contact (Thousand Oaks). 2024 Aug 13;7:25152564241273646. doi: 10.1177/25152564241273646. eCollection 2024 Jan-Dec. Contact (Thousand Oaks). 2024. PMID: 39139576 Free PMC article. Review.
-
Tex2 is required for lysosomal functions at TMEM55-dependent ER membrane contact sites.J Cell Biol. 2023 Apr 3;222(4):e202205133. doi: 10.1083/jcb.202205133. Epub 2023 Jan 27. J Cell Biol. 2023. PMID: 36705603 Free PMC article.
-
Transmembrane Membrane Readers form a Novel Class of Proteins That Include Peripheral Phosphoinositide Recognition Domains and Viral Spikes.Membranes (Basel). 2022 Nov 17;12(11):1161. doi: 10.3390/membranes12111161. Membranes (Basel). 2022. PMID: 36422153 Free PMC article. Review.
-
PDZD8-deficient mice manifest behavioral abnormalities related to emotion, cognition, and adaptation due to dyslipidemia in the brain.Mol Brain. 2023 Jan 19;16(1):11. doi: 10.1186/s13041-023-01002-4. Mol Brain. 2023. PMID: 36658656 Free PMC article.
-
Lipid Polarization during Cytokinesis.Cells. 2022 Dec 8;11(24):3977. doi: 10.3390/cells11243977. Cells. 2022. PMID: 36552741 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
