

Phase separation of a plant virus movement protein and cellular factors support virus-host interactions
- PMID: 34543360
- PMCID: PMC8483311
- DOI: 10.1371/journal.ppat.1009622
Phase separation of a plant virus movement protein and cellular factors support virus-host interactions
Abstract
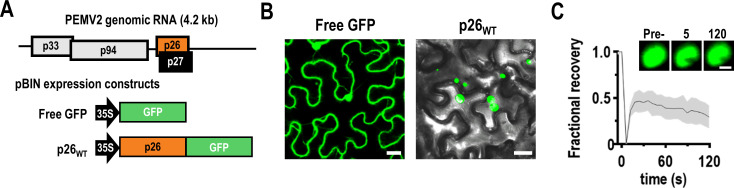
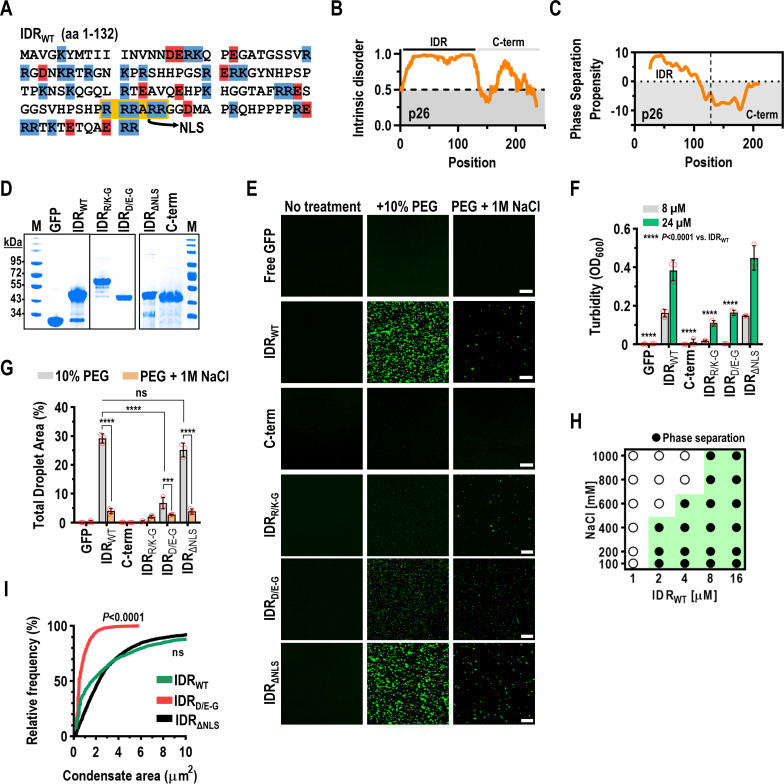
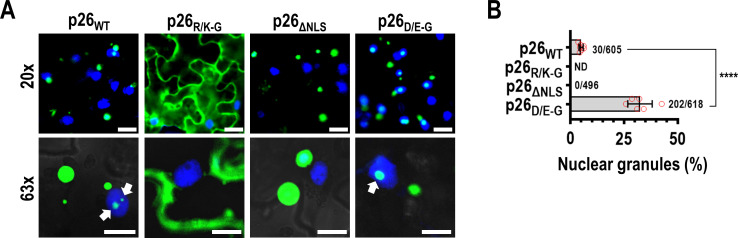
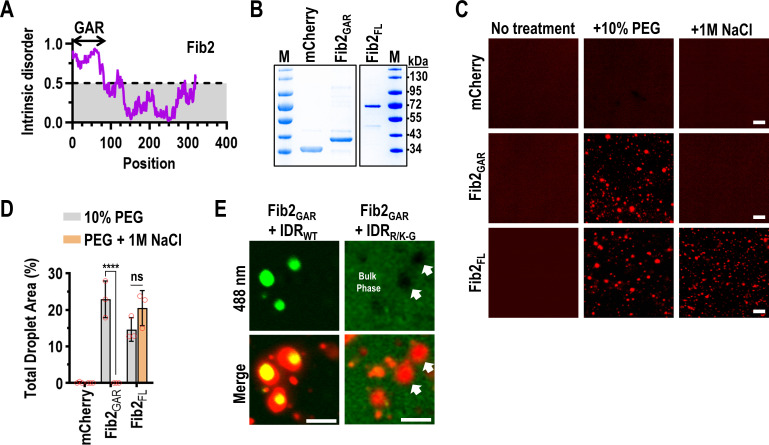
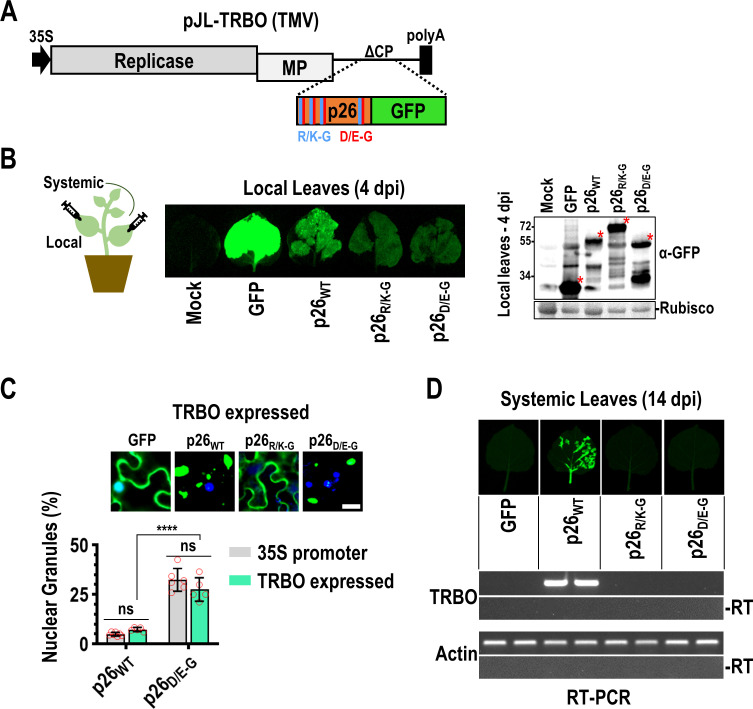
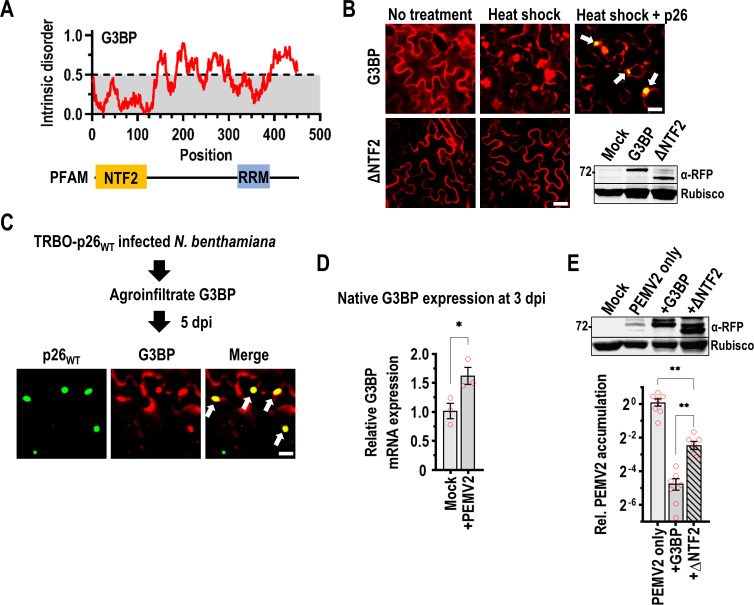
Both cellular and viral proteins can undergo phase separation and form membraneless compartments that concentrate biomolecules. The p26 movement protein from single-stranded, positive-sense Pea enation mosaic virus 2 (PEMV2) separates into a dense phase in nucleoli where p26 and related orthologues must interact with fibrillarin (Fib2) as a pre-requisite for systemic virus movement. Using in vitro assays, viral ribonucleoprotein complexes containing p26, Fib2, and PEMV2 genomic RNAs formed droplets that may provide the basis for self-assembly in planta. Mutating basic p26 residues (R/K-G) blocked droplet formation and partitioning into Fib2 droplets or the nucleolus and prevented systemic movement of a Tobacco mosaic virus (TMV) vector in Nicotiana benthamiana. Mutating acidic residues (D/E-G) reduced droplet formation in vitro, increased nucleolar retention 6.5-fold, and prevented systemic movement of TMV, thus demonstrating that p26 requires electrostatic interactions for droplet formation and charged residues are critical for nucleolar trafficking and virus movement. p26 readily partitioned into stress granules (SGs), which are membraneless compartments that assemble by clustering of the RNA binding protein G3BP following stress. G3BP is upregulated during PEMV2 infection and over-expression of G3BP restricted PEMV2 RNA accumulation >20-fold. Deletion of the NTF2 domain that is required for G3BP condensation restored PEMV2 RNA accumulation >4-fold, demonstrating that phase separation enhances G3BP antiviral activity. These results indicate that p26 partitions into membraneless compartments with either proviral (Fib2) or antiviral (G3BP) factors.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

Similar articles
-
The Multifunctional Long-Distance Movement Protein of Pea Enation Mosaic Virus 2 Protects Viral and Host Transcripts from Nonsense-Mediated Decay.mBio. 2020 Mar 10;11(2):e00204-20. doi: 10.1128/mBio.00204-20. mBio. 2020. PMID: 32156817 Free PMC article.
-
Hijacking of the nucleolar protein fibrillarin by TGB1 is required for cell-to-cell movement of Barley stripe mosaic virus.Mol Plant Pathol. 2018 May;19(5):1222-1237. doi: 10.1111/mpp.12612. Epub 2017 Dec 18. Mol Plant Pathol. 2018. PMID: 28872759 Free PMC article.
-
A Distinct, Non-Virion Plant Virus Movement Protein Encoded by a Crinivirus Essential for Systemic Infection.mBio. 2018 Nov 20;9(6):e02230-18. doi: 10.1128/mBio.02230-18. mBio. 2018. PMID: 30459200 Free PMC article.
-
Plant virus replication and movement.Virology. 2015 May;479-480:657-71. doi: 10.1016/j.virol.2015.01.025. Epub 2015 Mar 3. Virology. 2015. PMID: 25746797 Review.
-
Replication of tobacco mosaic virus RNA.Philos Trans R Soc Lond B Biol Sci. 1999 Mar 29;354(1383):613-27. doi: 10.1098/rstb.1999.0413. Philos Trans R Soc Lond B Biol Sci. 1999. PMID: 10212941 Free PMC article. Review.
Cited by
-
Cell Fractionation and the Identification of Host Proteins Involved in Plant-Virus Interactions.Pathogens. 2024 Jan 5;13(1):53. doi: 10.3390/pathogens13010053. Pathogens. 2024. PMID: 38251360 Free PMC article. Review.
-
Unraveling the Mechanisms of Virus-Induced Symptom Development in Plants.Plants (Basel). 2023 Jul 31;12(15):2830. doi: 10.3390/plants12152830. Plants (Basel). 2023. PMID: 37570983 Free PMC article. Review.
-
Subversion of selective autophagy for the biogenesis of tombusvirus replication organelles inhibits autophagy.PLoS Pathog. 2024 Mar 14;20(3):e1012085. doi: 10.1371/journal.ppat.1012085. eCollection 2024 Mar. PLoS Pathog. 2024. PMID: 38484009 Free PMC article.
-
Liaisons dangereuses: Intrinsic Disorder in Cellular Proteins Recruited to Viral Infection-Related Biocondensates.Int J Mol Sci. 2023 Jan 21;24(3):2151. doi: 10.3390/ijms24032151. Int J Mol Sci. 2023. PMID: 36768473 Free PMC article.
-
P1 of turnip mosaic virus interacts with NOD19 for vigorous infection.Front Microbiol. 2023 Jun 23;14:1216950. doi: 10.3389/fmicb.2023.1216950. eCollection 2023. Front Microbiol. 2023. PMID: 37426031 Free PMC article.
References
-
- Dolgin E. What lava lamps and vinaigrette can teach us about cell biology. Nature. 2018;555(7696):300–2. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
