

Establishment, maintenance, and recall of inflammatory memory
- PMID: 34320411
- PMCID: PMC8500942
- DOI: 10.1016/j.stem.2021.07.001
Establishment, maintenance, and recall of inflammatory memory
Abstract
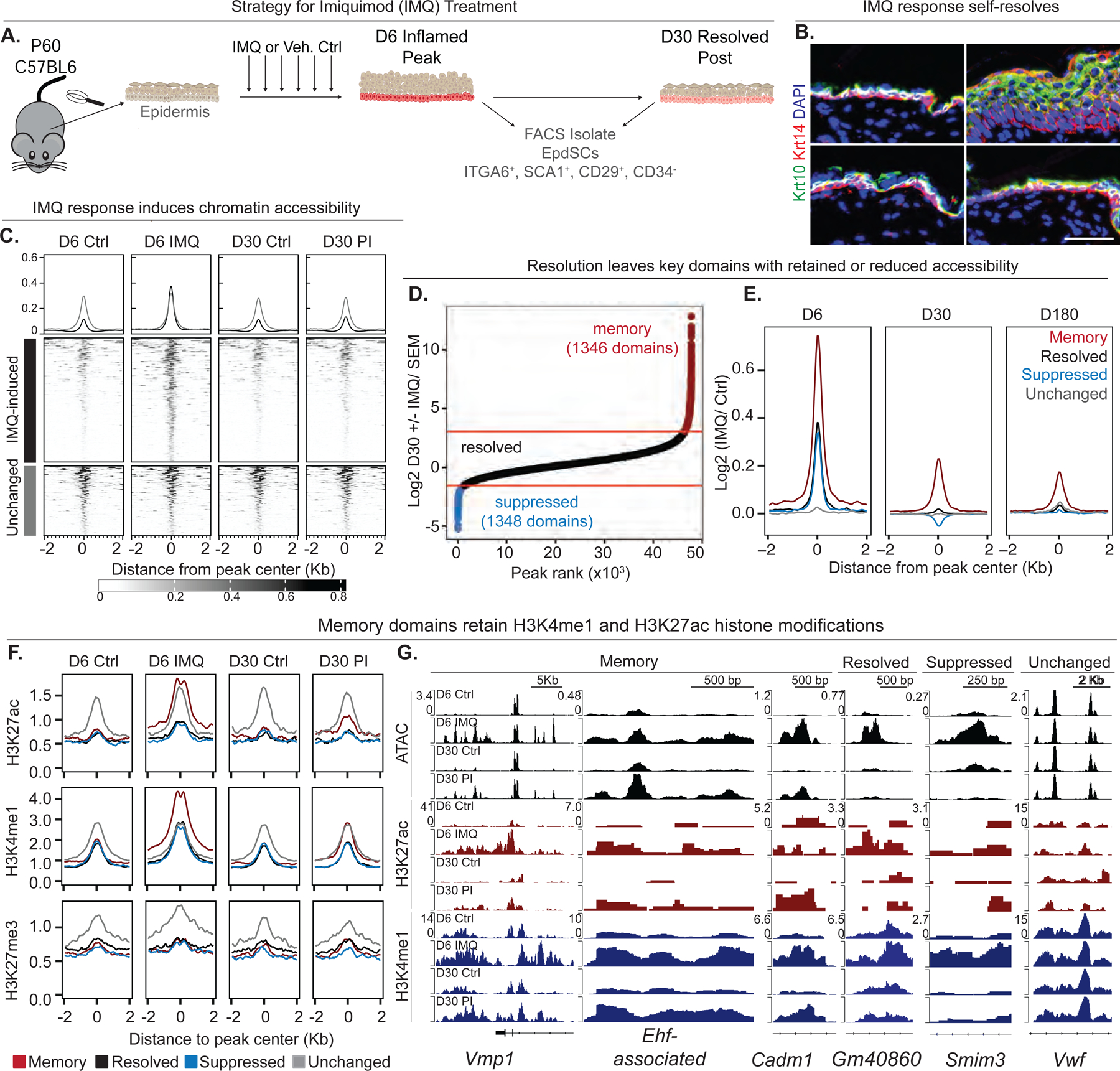
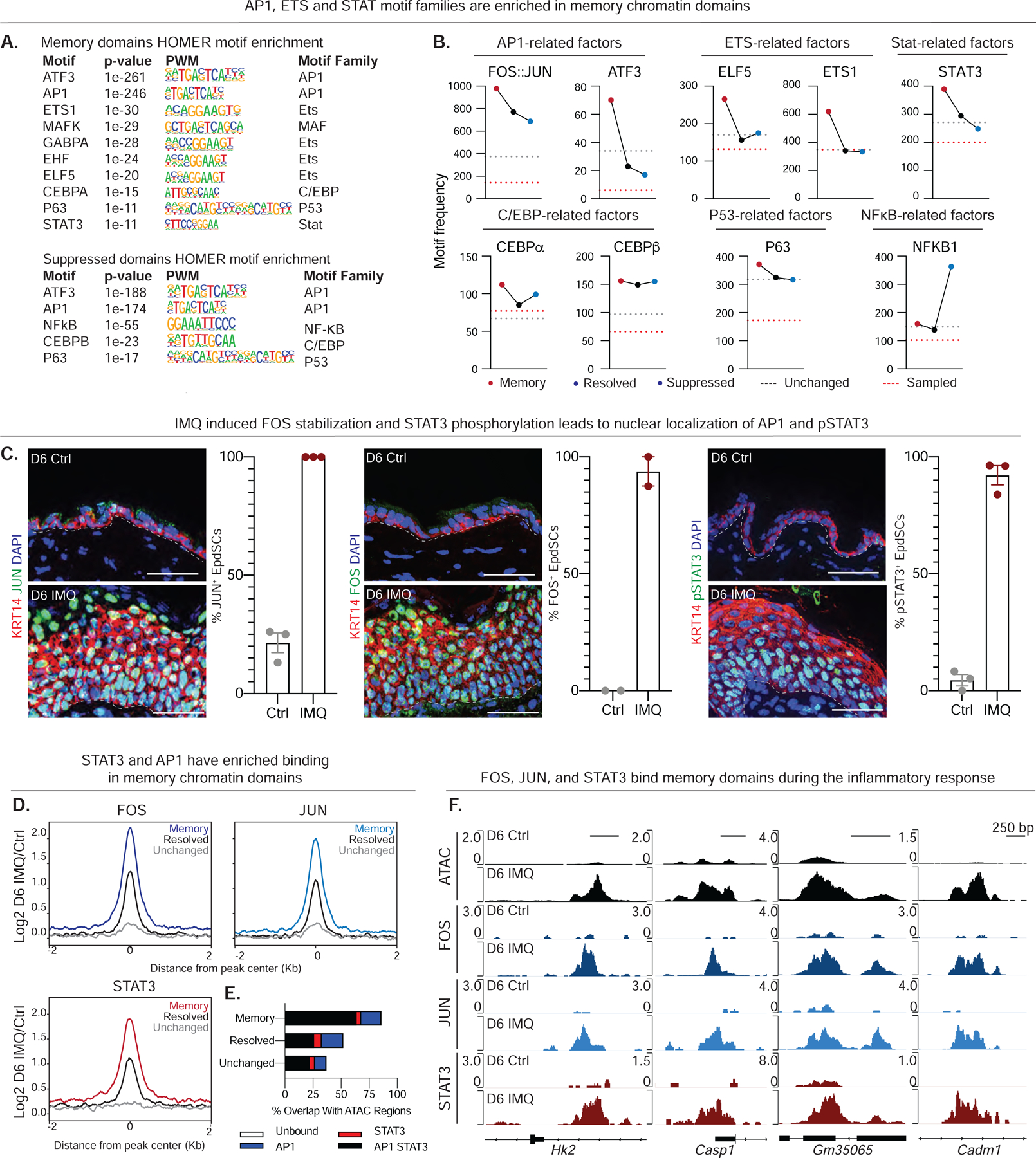
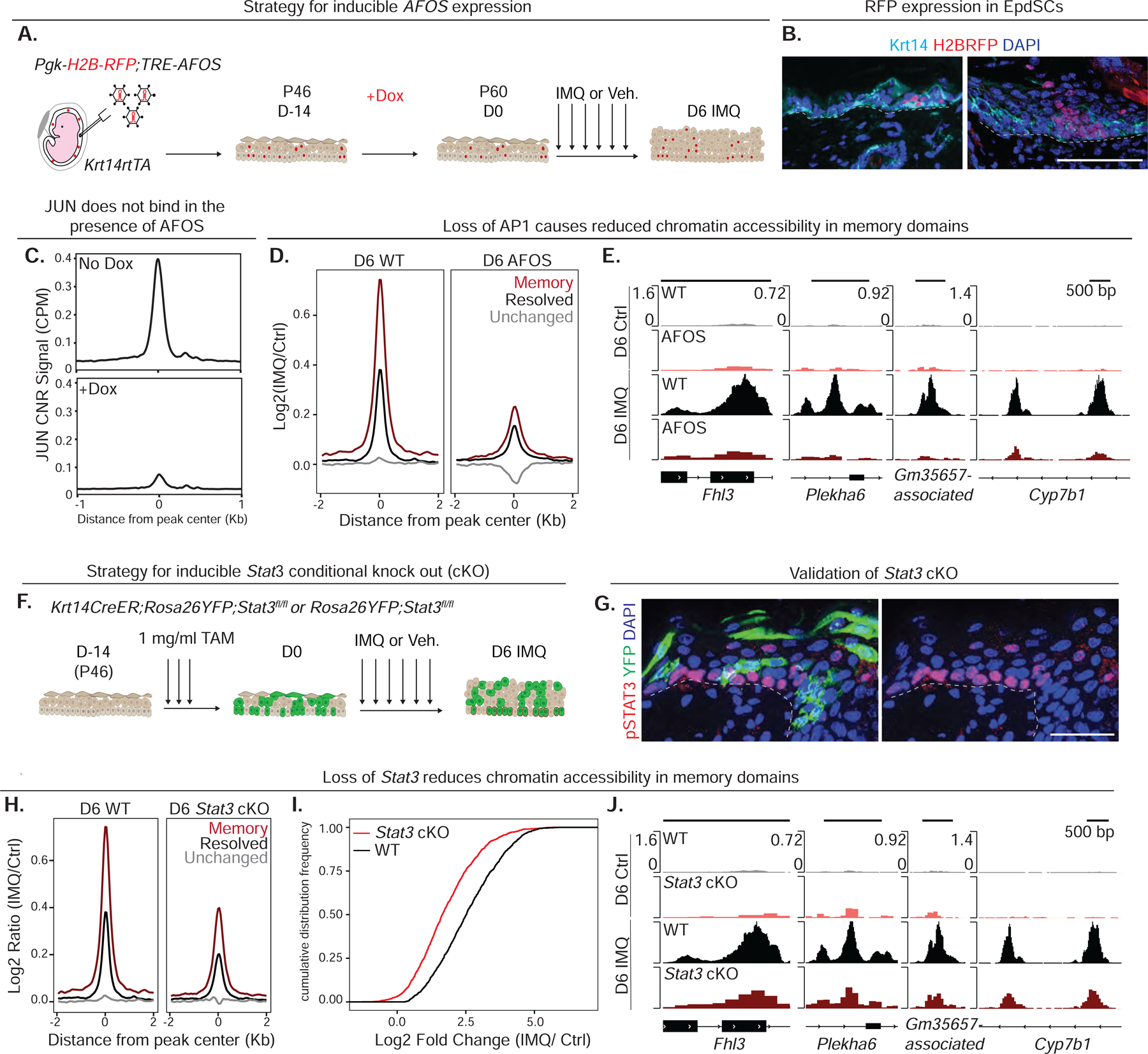
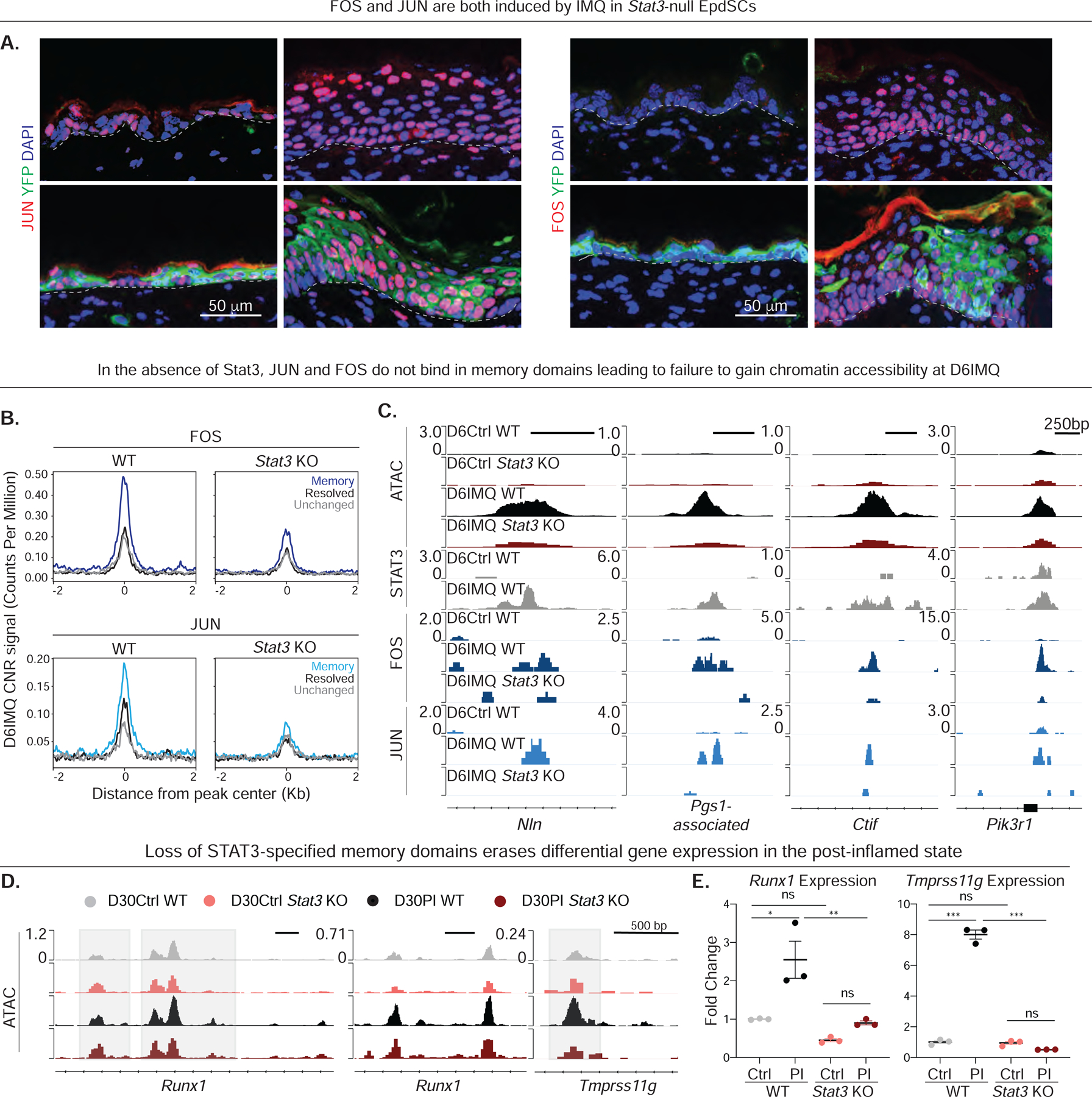
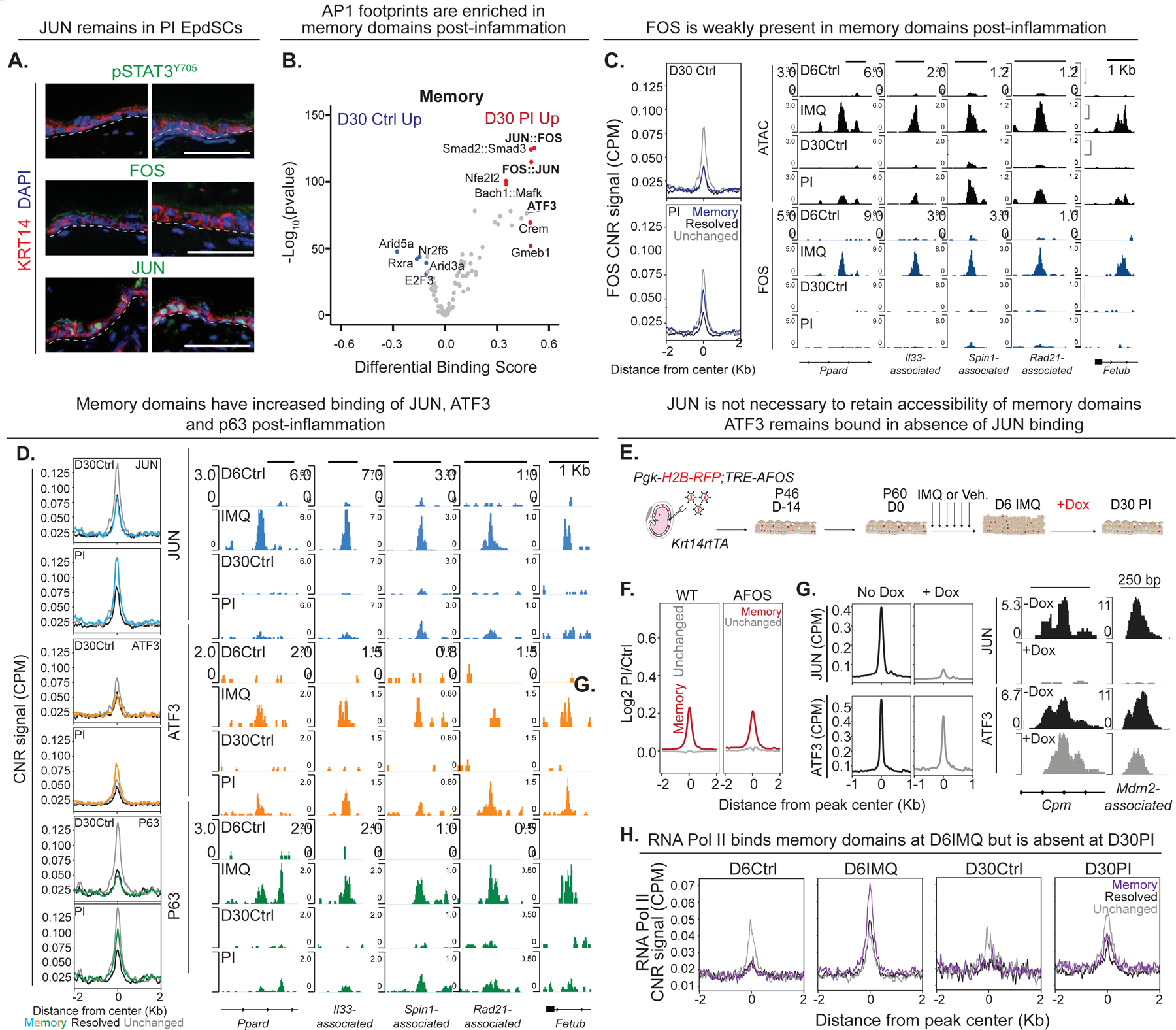
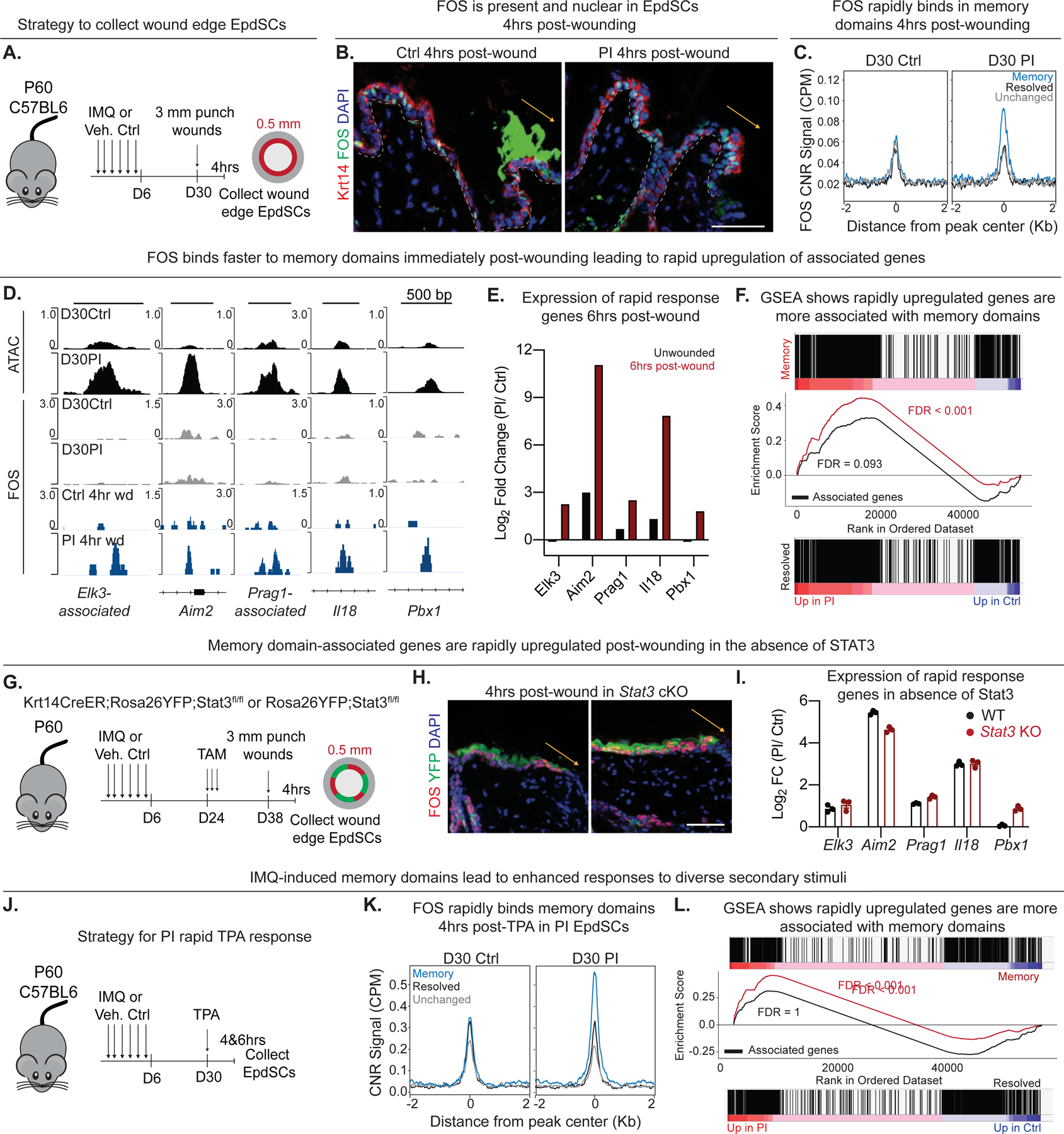
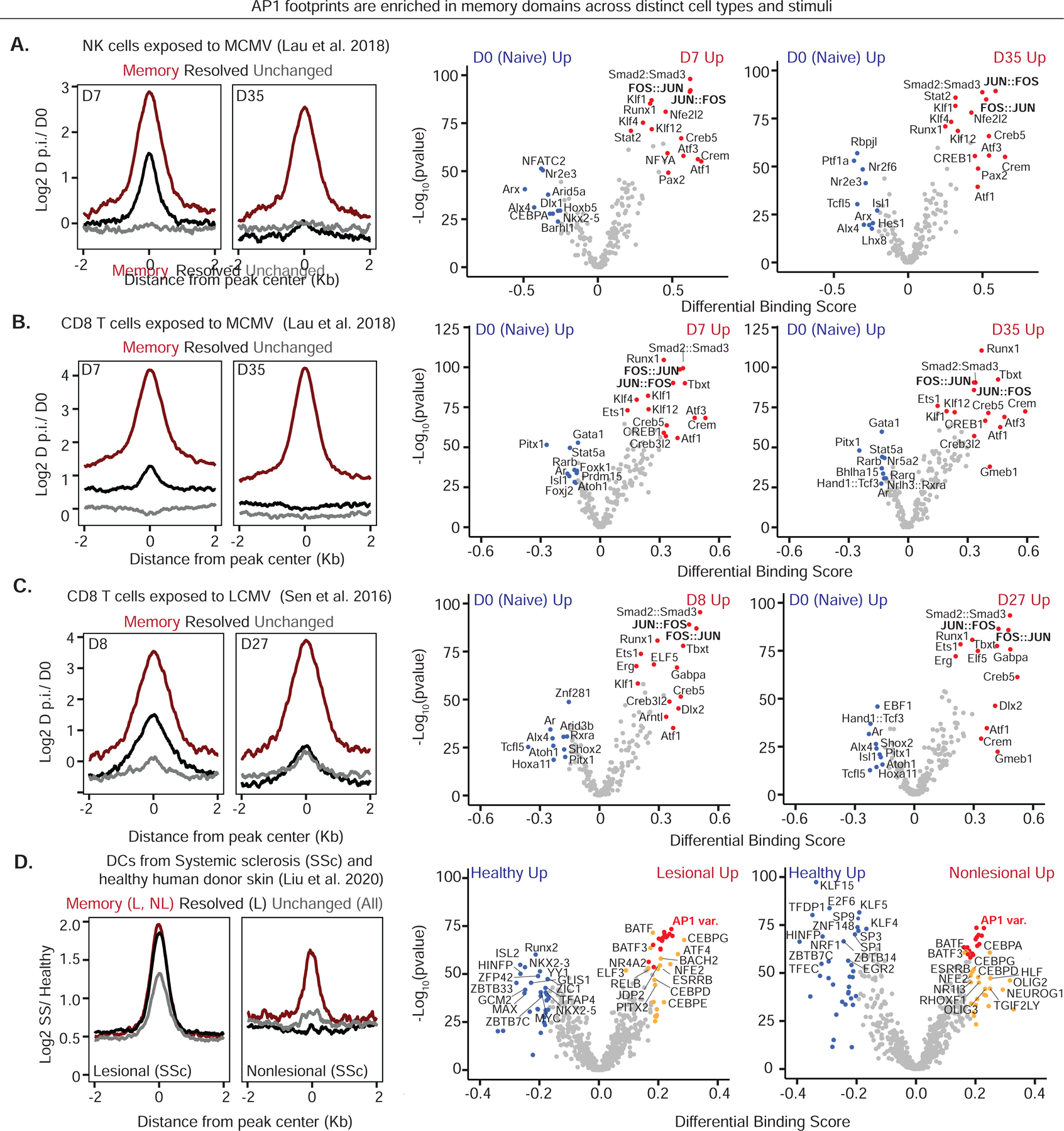
Known for nearly a century but through mechanisms that remain elusive, cells retain a memory of inflammation that equips them to react quickly and broadly to diverse secondary stimuli. Using murine epidermal stem cells as a model, we elucidate how cells establish, maintain, and recall inflammatory memory. Specifically, we landscape and functionally interrogate temporal, dynamic changes to chromatin accessibility, histone modifications, and transcription factor binding that occur during inflammation, post-resolution, and in memory recall following injury. We unearth an essential, unifying role for the general stress-responsive transcription factor FOS, which partners with JUN and cooperates with stimulus-specific STAT3 to establish memory; JUN then remains with other homeostatic factors on memory domains, facilitating rapid FOS re-recruitment and gene re-activation upon diverse secondary challenges. Extending our findings, we offer a comprehensive, potentially universal mechanism behind inflammatory memory and less discriminate recall phenomena with profound implications for tissue fitness in health and disease.
Keywords: AP1 transcription factors; ATAC sequencing; CUT&RUN; ChIP sequencing; FOS; FOS:JUN; STAT3; broadened immune protection; epigenetic memory; histone modifications; inflammation; inflammatory disorders; inflammatory memory; tissue stem cells; trained immunity.
Copyright © 2021 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing financial interests. E.F. serves as an advisory board member for Cell Stem Cell and serves on the scientific advisory boards of L’Oreal and Arsenal Biosciences.
Figures
Comment in
-
Stress-responsive transcription factors train stem cells to remember.Cell Stem Cell. 2021 Oct 7;28(10):1679-1680. doi: 10.1016/j.stem.2021.09.005. Cell Stem Cell. 2021. PMID: 34624227
Similar articles
-
The Role of Epigenetic Regulation in Transcriptional Memory in the Immune System.Adv Protein Chem Struct Biol. 2017;106:43-69. doi: 10.1016/bs.apcsb.2016.09.002. Epub 2016 Oct 8. Adv Protein Chem Struct Biol. 2017. PMID: 28057215 Review.
-
Stress-responsive transcription factors train stem cells to remember.Cell Stem Cell. 2021 Oct 7;28(10):1679-1680. doi: 10.1016/j.stem.2021.09.005. Cell Stem Cell. 2021. PMID: 34624227
-
A Transcription Factor Pulse Can Prime Chromatin for Heritable Transcriptional Memory.Mol Cell Biol. 2017 Feb 1;37(4):e00372-16. doi: 10.1128/MCB.00372-16. Print 2017 Feb 15. Mol Cell Biol. 2017. PMID: 27920256 Free PMC article.
-
Nuclear PKC-θ facilitates rapid transcriptional responses in human memory CD4+ T cells through p65 and H2B phosphorylation.J Cell Sci. 2016 Jun 15;129(12):2448-61. doi: 10.1242/jcs.181248. Epub 2016 May 5. J Cell Sci. 2016. PMID: 27149922 Free PMC article.
-
Epigenetic landscape of amphetamine and methamphetamine addiction in rodents.Epigenetics. 2015;10(7):574-80. doi: 10.1080/15592294.2015.1055441. Epigenetics. 2015. PMID: 26023847 Free PMC article. Review.
Cited by
-
Key platelet genes play important roles in predicting the prognosis of sepsis.Sci Rep. 2024 Oct 9;14(1):23530. doi: 10.1038/s41598-024-74052-w. Sci Rep. 2024. PMID: 39384856 Free PMC article.
-
Stem cells expand potency and alter tissue fitness by accumulating diverse epigenetic memories.Science. 2021 Nov 26;374(6571):eabh2444. doi: 10.1126/science.abh2444. Epub 2021 Nov 26. Science. 2021. PMID: 34822296 Free PMC article.
-
Healing Ability of Central Corneal Epithelium in Rabbit Ocular Surface Injury Models.Transl Vis Sci Technol. 2022 Jun 1;11(6):28. doi: 10.1167/tvst.11.6.28. Transl Vis Sci Technol. 2022. PMID: 35771535 Free PMC article.
-
Maternal diet alters long-term innate immune cell memory in fetal and juvenile hematopoietic stem and progenitor cells in nonhuman primate offspring.Cell Rep. 2023 Apr 25;42(4):112393. doi: 10.1016/j.celrep.2023.112393. Epub 2023 Apr 13. Cell Rep. 2023. PMID: 37058409 Free PMC article.
-
Epigenetic memory of radiotherapy in dermal fibroblasts impairs wound repair capacity in cancer survivors.Nat Commun. 2024 Oct 28;15(1):9286. doi: 10.1038/s41467-024-53295-1. Nat Commun. 2024. PMID: 39468077 Free PMC article.
References
-
- Adam RC, Yang H, Ge Y, Infarinato NR, Gur-Cohen S, Miao Y, Wang P, Zhao Y, Lu CP, Kim JE, et al. (2020). NFI transcription factors provide chromatin access to maintain stem cell identity while preventing unintended lineage fate choices. Nat Cell Biol 22, 640–650. 10.1038/s41556-020-0513-0. - DOI - PMC - PubMed
-
- Adam RC, Yang H, Ge Y, Lien WH, Wang P, Zhao Y, Polak L, Levorse J, Baksh SC, Zheng D, and Fuchs E (2018). Temporal Layering of Signaling Effectors Drives Chromatin Remodeling during Hair Follicle Stem Cell Lineage Progression. Cell Stem Cell 22, 398–413.e397. 10.1016/j.stem.2017.12.004. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
