

eIF4E3 forms an active eIF4F complex during stresses (eIF4FS) targeting mTOR and re-programs the translatome
- PMID: 33893802
- PMCID: PMC8136781
- DOI: 10.1093/nar/gkab267
eIF4E3 forms an active eIF4F complex during stresses (eIF4FS) targeting mTOR and re-programs the translatome
Abstract
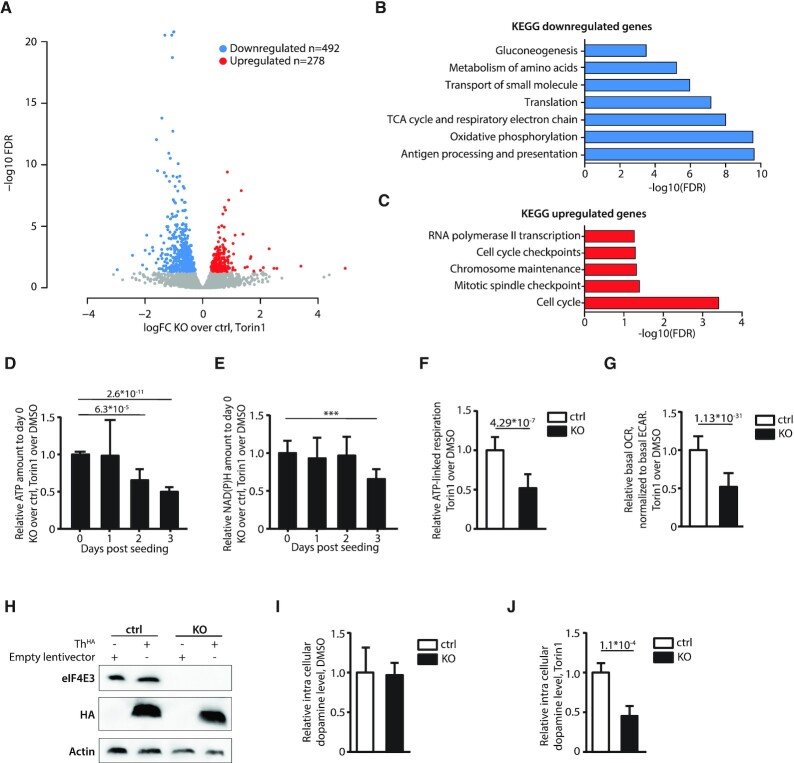
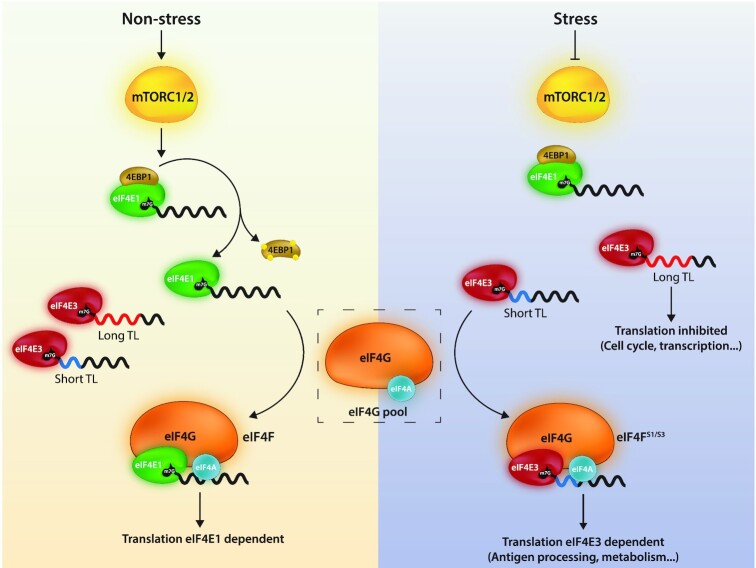
The eIF4E are a family of initiation factors that bind the mRNA 5' cap, regulating the proteome and the cellular phenotype. eIF4E1 mediates global translation and its activity is controlled via the PI3K/AKT/mTOR pathway. mTOR down-regulation results in eIF4E1 sequestration into an inactive complex with the 4E binding proteins (4EBPs). The second member, eIF4E2, regulates the translatome during hypoxia. However, the exact function of the third member, eIF4E3, has remained elusive. We have dissected its function using a range of techniques. Starting from the observation that it does not interact with 4EBP1, we demonstrate that eIF4E3 recruitment into an eIF4F complex occurs when Torin1 inhibits the mTOR pathway. Ribo-seq studies demonstrate that this complex (eIF4FS) is translationally active during stress and that it selects specific mRNA populations based on 5' TL (UTR) length. The interactome reveals that it associates with cellular proteins beyond the cognate initiation factors, suggesting that it may have 'moon-lighting' functions. Finally, we provide evidence that cellular metabolism is altered in an eIF4E3 KO background but only upon Torin1 treatment. We propose that eIF4E3 acts as a second branch of the integrated stress response, re-programming the translatome to promote 'stress resistance' and adaptation.
© The Author(s) 2021. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
Similar articles
-
Distinct recruitment of human eIF4E isoforms to processing bodies and stress granules.BMC Mol Biol. 2016 Aug 30;17(1):21. doi: 10.1186/s12867-016-0072-x. BMC Mol Biol. 2016. PMID: 27578149 Free PMC article.
-
Translation initiation complex eIF4F is a therapeutic target for dual mTOR kinase inhibitors in non-Hodgkin lymphoma.Oncotarget. 2015 Apr 20;6(11):9488-501. doi: 10.18632/oncotarget.3378. Oncotarget. 2015. PMID: 25839159 Free PMC article.
-
Hepatitis C virus NS5A binds to the mRNA cap-binding eukaryotic translation initiation 4F (eIF4F) complex and up-regulates host translation initiation machinery through eIF4E-binding protein 1 inactivation.J Biol Chem. 2012 Feb 10;287(7):5042-58. doi: 10.1074/jbc.M111.308916. Epub 2011 Dec 19. J Biol Chem. 2012. PMID: 22184107 Free PMC article.
-
A Cap for Every Occasion: Alternative eIF4F Complexes.Trends Biochem Sci. 2016 Oct;41(10):821-823. doi: 10.1016/j.tibs.2016.05.009. Epub 2016 Jun 6. Trends Biochem Sci. 2016. PMID: 27283511 Free PMC article. Review.
-
Manipulation of the host translation initiation complex eIF4F by DNA viruses.Biochem Soc Trans. 2010 Dec;38(6):1511-6. doi: 10.1042/BST0381511. Biochem Soc Trans. 2010. PMID: 21118117 Review.
Cited by
-
eIF4E-independent translation is largely eIF3d-dependent.Nat Commun. 2024 Aug 6;15(1):6692. doi: 10.1038/s41467-024-51027-z. Nat Commun. 2024. PMID: 39107322 Free PMC article.
-
eIF4E1b is a non-canonical eIF4E protecting maternal dormant mRNAs.EMBO Rep. 2024 Jan;25(1):404-427. doi: 10.1038/s44319-023-00006-4. Epub 2023 Dec 14. EMBO Rep. 2024. PMID: 38177902 Free PMC article.
-
RNA modifications in cancer.Br J Cancer. 2023 Aug;129(2):204-221. doi: 10.1038/s41416-023-02275-1. Epub 2023 Apr 24. Br J Cancer. 2023. PMID: 37095185 Free PMC article. Review.
-
Non-canonical mRNA translation initiation in cell stress and cancer.NAR Cancer. 2024 May 31;6(2):zcae026. doi: 10.1093/narcan/zcae026. eCollection 2024 Jun. NAR Cancer. 2024. PMID: 38828390 Free PMC article. Review.
-
eIF4EHP promotes Ldh mRNA translation in and fruit fly adaptation to hypoxia.EMBO Rep. 2023 Jul 5;24(7):e56460. doi: 10.15252/embr.202256460. Epub 2023 May 5. EMBO Rep. 2023. PMID: 37144276 Free PMC article.
References
-
- Pannevis M.C., Houlihan D.F.. The energetic cost of protein synthesis in isolated hepatocytes of rainbow trout (Oncorhynchus mykiss). J. Comp. Physiol. [B]. 1992; 162:393–400. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
