

Conformational interconversion of MLKL and disengagement from RIPK3 precede cell death by necroptosis
- PMID: 33850121
- PMCID: PMC8044208
- DOI: 10.1038/s41467-021-22400-z
Conformational interconversion of MLKL and disengagement from RIPK3 precede cell death by necroptosis
Abstract
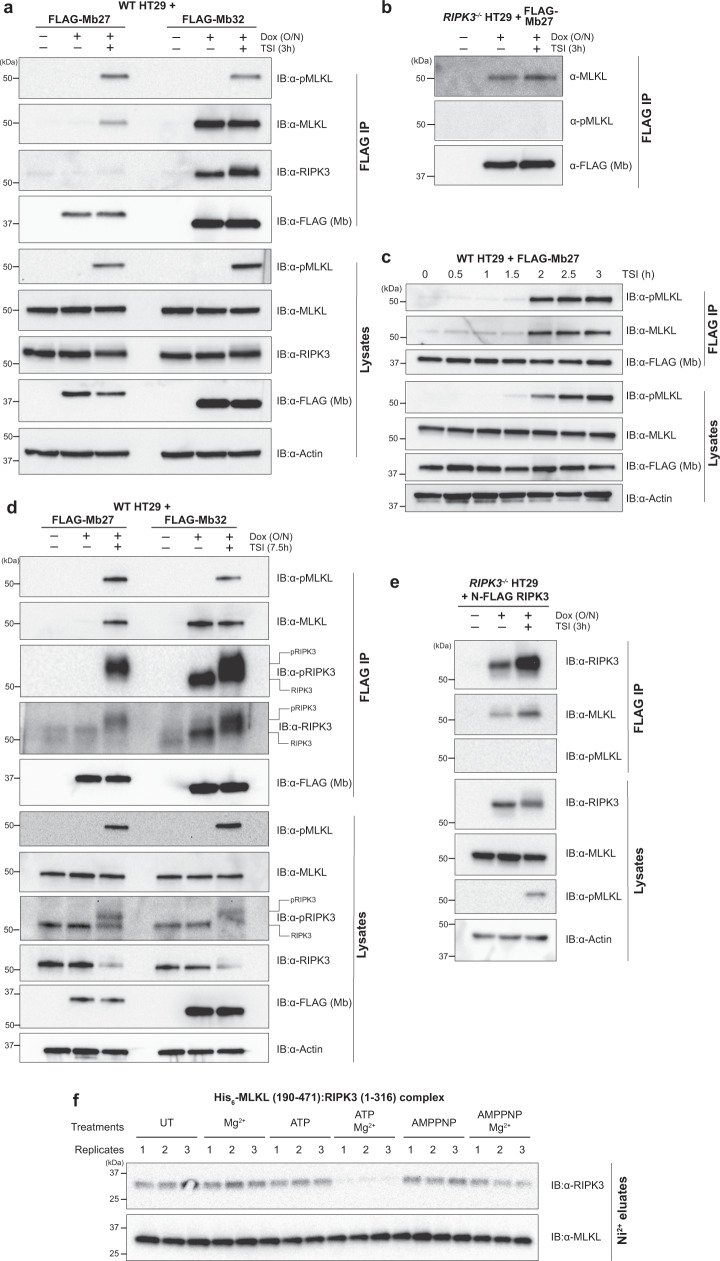
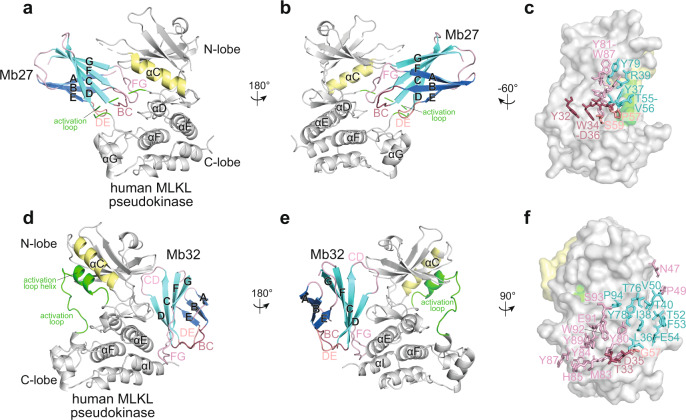
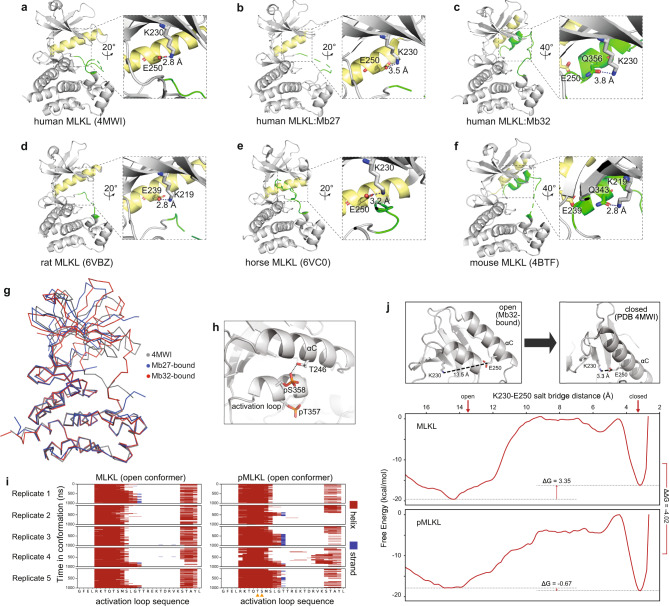
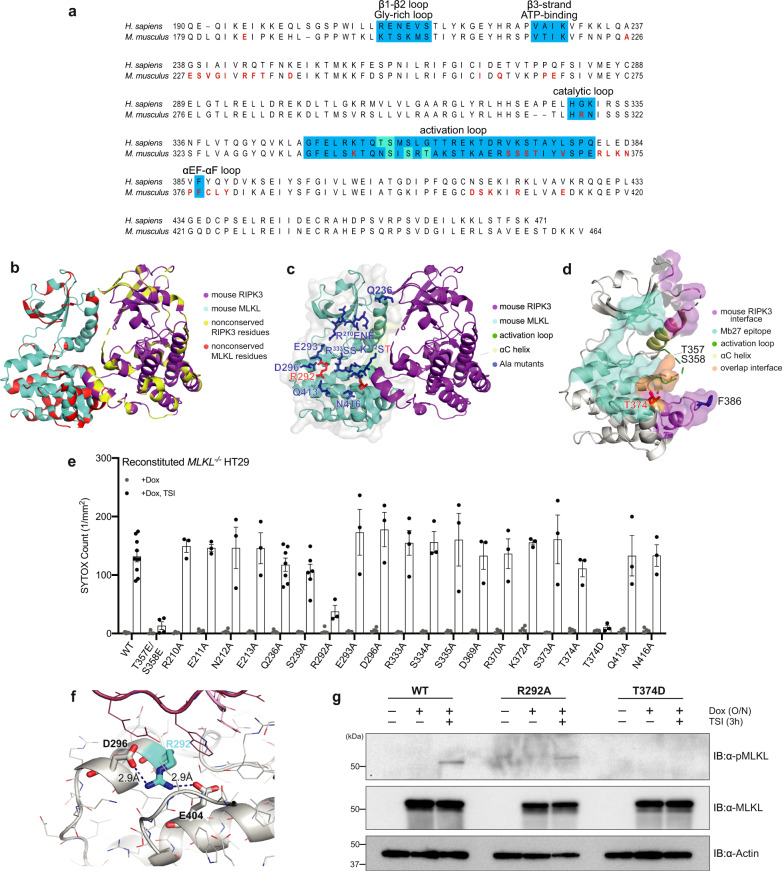
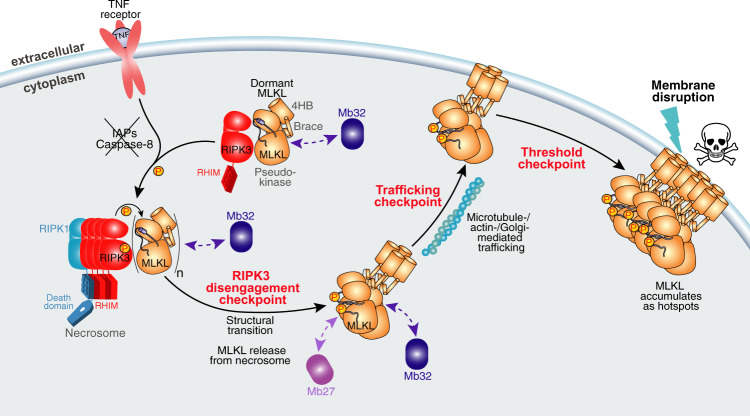
Phosphorylation of the MLKL pseudokinase by the RIPK3 kinase leads to MLKL oligomerization, translocation to, and permeabilization of, the plasma membrane to induce necroptotic cell death. The precise choreography of MLKL activation remains incompletely understood. Here, we report Monobodies, synthetic binding proteins, that bind the pseudokinase domain of MLKL within human cells and their crystal structures in complex with the human MLKL pseudokinase domain. While Monobody-32 constitutively binds the MLKL hinge region, Monobody-27 binds MLKL via an epitope that overlaps the RIPK3 binding site and is only exposed after phosphorylated MLKL disengages from RIPK3 following necroptotic stimulation. The crystal structures identified two distinct conformations of the MLKL pseudokinase domain, supporting the idea that a conformational transition accompanies MLKL disengagement from RIPK3. These studies provide further evidence that MLKL undergoes a large conformational change upon activation, and identify MLKL disengagement from RIPK3 as a key regulatory step in the necroptosis pathway.
Conflict of interest statement
S.E.G., A.L.S., C.F., S.N.Y., E.J.P., J.M.H., P.E.C., and J.M.M. contribute, or have contributed, to a project developing necroptosis inhibitors in collaboration with Anaxis Pharma Pty Ltd. A.K. and S.K. are listed as inventors on issued and pending patents on the Monobody technology filed by The University of Chicago (US Patent 9512199 B2 and related pending applications). The other authors declare no competing interests.
Figures
Similar articles
-
Human RIPK3 maintains MLKL in an inactive conformation prior to cell death by necroptosis.Nat Commun. 2021 Nov 22;12(1):6783. doi: 10.1038/s41467-021-27032-x. Nat Commun. 2021. PMID: 34811356 Free PMC article.
-
Identification of MLKL membrane translocation as a checkpoint in necroptotic cell death using Monobodies.Proc Natl Acad Sci U S A. 2020 Apr 14;117(15):8468-8475. doi: 10.1073/pnas.1919960117. Epub 2020 Mar 31. Proc Natl Acad Sci U S A. 2020. PMID: 32234780 Free PMC article.
-
Necroptosis signalling is tuned by phosphorylation of MLKL residues outside the pseudokinase domain activation loop.Biochem J. 2015 Oct 15;471(2):255-65. doi: 10.1042/BJ20150678. Epub 2015 Aug 17. Biochem J. 2015. PMID: 26283547
-
The regulation of necroptosis by post-translational modifications.Cell Death Differ. 2021 Mar;28(3):861-883. doi: 10.1038/s41418-020-00722-7. Epub 2021 Jan 18. Cell Death Differ. 2021. PMID: 33462412 Free PMC article. Review.
-
Surviving death: emerging concepts of RIPK3 and MLKL ubiquitination in the regulation of necroptosis.FEBS J. 2023 Jan;290(1):37-54. doi: 10.1111/febs.16255. Epub 2021 Nov 16. FEBS J. 2023. PMID: 34710282 Review.
Cited by
-
The intracellular domains of the EphB6 and EphA10 receptor tyrosine pseudokinases function as dynamic signalling hubs.Biochem J. 2021 Sep 17;478(17):3351-3371. doi: 10.1042/BCJ20210572. Biochem J. 2021. PMID: 34431498 Free PMC article.
-
Necroptosis at a glance.J Cell Sci. 2022 Sep 1;135(17):jcs260091. doi: 10.1242/jcs.260091. Epub 2022 Sep 13. J Cell Sci. 2022. PMID: 36098620 Free PMC article.
-
There's more to death than life: Noncatalytic functions in kinase and pseudokinase signaling.J Biol Chem. 2021 Jan-Jun;296:100705. doi: 10.1016/j.jbc.2021.100705. Epub 2021 Apr 22. J Biol Chem. 2021. PMID: 33895136 Free PMC article. Review.
-
Quantitative phosphoproteomic analysis reveals unique cAMP signaling pools emanating from AC2 and AC6 in human airway smooth muscle cells.Front Physiol. 2023 Feb 28;14:1149063. doi: 10.3389/fphys.2023.1149063. eCollection 2023. Front Physiol. 2023. PMID: 36926196 Free PMC article.
-
The VEGFR/PDGFR tyrosine kinase inhibitor, ABT-869, blocks necroptosis by targeting RIPK1 kinase.Biochem J. 2023 May 15;480(9):665-684. doi: 10.1042/BCJ20230035. Biochem J. 2023. PMID: 37115711 Free PMC article.
References
-
- He S, et al. Receptor interacting protein kinase-3 determines cellular necrotic response to TNF-alpha. Cell. 2009;137:1100–1111. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
