

Novel Asaia bogorensis Signal Sequences for Plasmodium Inhibition in Anopheles stephensi
- PMID: 33664722
- PMCID: PMC7921796
- DOI: 10.3389/fmicb.2021.633667
Novel Asaia bogorensis Signal Sequences for Plasmodium Inhibition in Anopheles stephensi
Abstract
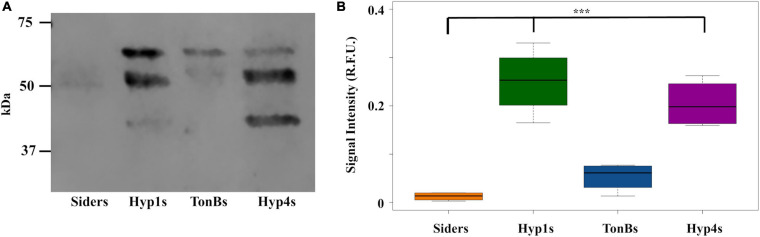
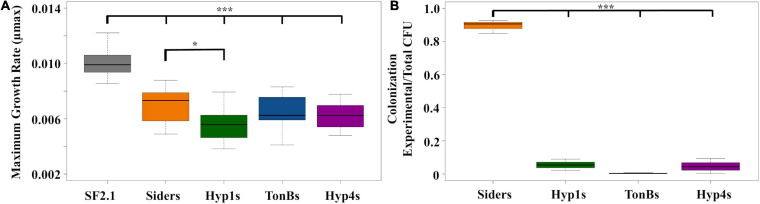
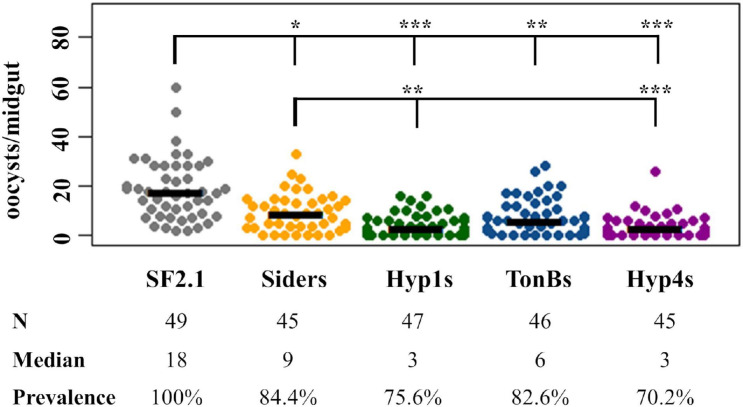
Mosquitoes vector many pathogens that cause human disease, such as malaria that is caused by parasites in the genus Plasmodium. Current strategies to control vector-transmitted diseases are hindered by mosquito and pathogen resistance, so research has turned to altering the microbiota of the vectors. In this strategy, called paratransgenesis, symbiotic bacteria are genetically modified to affect the mosquito's phenotype by engineering them to deliver antiplasmodial effector molecules into the midgut to kill parasites. One paratransgenesis candidate is Asaia bogorensis, a Gram-negative, rod-shaped bacterium colonizing the midgut, ovaries, and salivary glands of Anopheles sp. mosquitoes. However, common secretion signals from E. coli and closely related species do not function in Asaia. Here, we report evaluation of 20 native Asaia N-terminal signal sequences predicted from bioinformatics for their ability to mediate increased levels of antiplasmodial effector molecules directed to the periplasm and ultimately outside the cell. We tested the hypothesis that by increasing the amount of antiplasmodials released from the cell we would also increase parasite killing power. We scanned the Asaia bogorensis SF2.1 genome to identify signal sequences from extra-cytoplasmic proteins and fused these to the reporter protein alkaline phosphatase. Six signals resulted in significant levels of protein released from the Asaia bacterium. Three signals were successfully used to drive the release of the antimicrobial peptide, scorpine. Further testing in mosquitoes demonstrated that these three Asaia strains were able to suppress the number of oocysts formed after a blood meal containing P. berghei to a significantly greater degree than wild-type Asaia, although prevalence was not decreased beyond levels obtained with a previously isolated siderophore receptor signal sequence. We interpret these results to indicate that there is a maximum level of suppression that can be achieved when the effectors are constitutively driven due to stress on the symbionts. This suggests that simply increasing the amount of antiplasmodial effector molecules in the midgut is insufficient to create superior paratransgenic bacterial strains and that symbiont fitness must be considered as well.
Keywords: Anopheles; Asaia; Plasmodium; malaria; paratransgenesis; secretion.
Copyright © 2021 Grogan, Bennett, Moore and Lampe.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
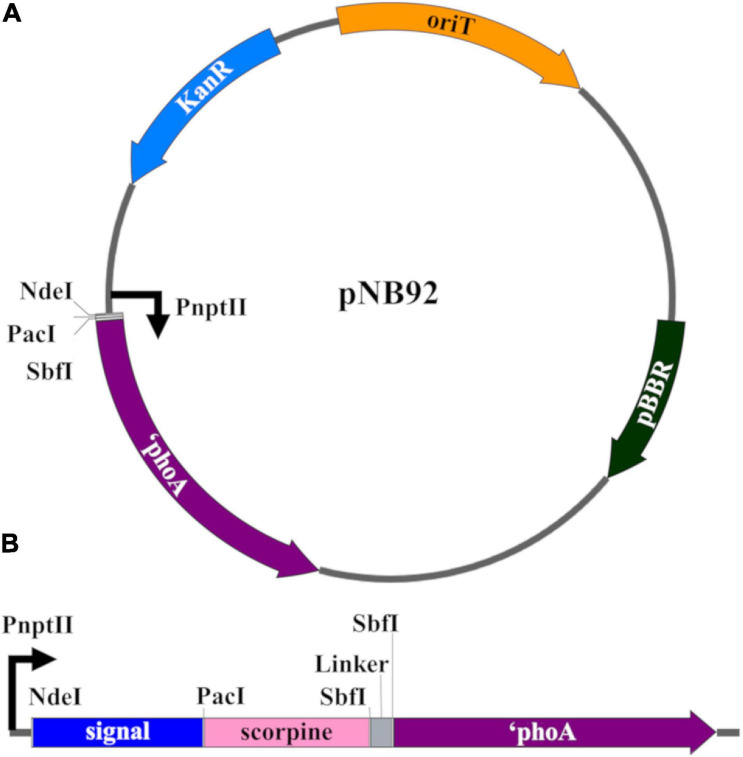
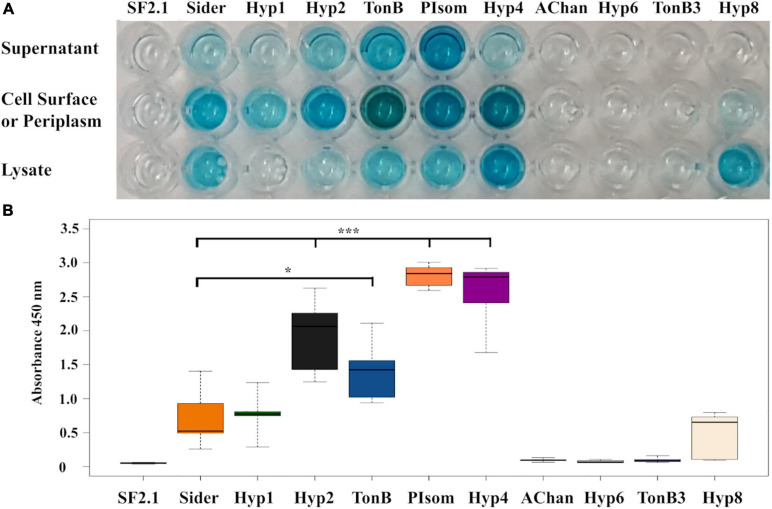
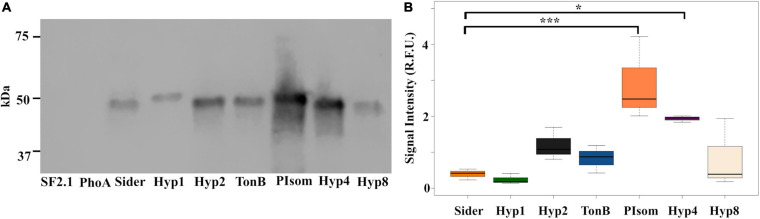
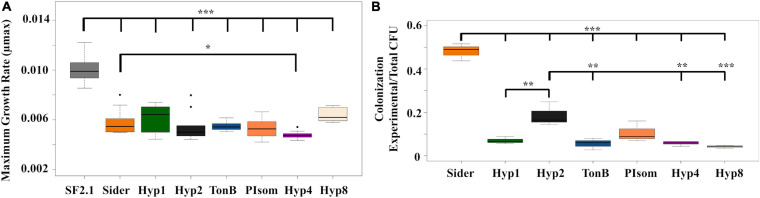
Similar articles
-
An evaluation of fusion partner proteins for paratransgenesis in Asaia bogorensis.PLoS One. 2022 Sep 1;17(9):e0273568. doi: 10.1371/journal.pone.0273568. eCollection 2022. PLoS One. 2022. PMID: 36048823 Free PMC article.
-
Inhibition of Plasmodium berghei Development in Mosquitoes by Effector Proteins Secreted from Asaia sp. Bacteria Using a Novel Native Secretion Signal.PLoS One. 2015 Dec 4;10(12):e0143541. doi: 10.1371/journal.pone.0143541. eCollection 2015. PLoS One. 2015. PMID: 26636338 Free PMC article.
-
Interactions between Asaia, Plasmodium and Anopheles: new insights into mosquito symbiosis and implications in malaria symbiotic control.Parasit Vectors. 2013 Jun 18;6(1):182. doi: 10.1186/1756-3305-6-182. Parasit Vectors. 2013. PMID: 23777746 Free PMC article.
-
Bacteria of the genus Asaia: a potential paratransgenic weapon against malaria.Adv Exp Med Biol. 2008;627:49-59. doi: 10.1007/978-0-387-78225-6_4. Adv Exp Med Biol. 2008. PMID: 18510013 Review.
-
Using bacteria to express and display anti-parasite molecules in mosquitoes: current and future strategies.Insect Biochem Mol Biol. 2005 Jul;35(7):699-707. doi: 10.1016/j.ibmb.2005.02.008. Epub 2005 Mar 25. Insect Biochem Mol Biol. 2005. PMID: 15894187 Review.
Cited by
-
The dynamic gut microbiota of zoophilic members of the Anopheles gambiae complex (Diptera: Culicidae).Sci Rep. 2022 Jan 27;12(1):1495. doi: 10.1038/s41598-022-05437-y. Sci Rep. 2022. PMID: 35087127 Free PMC article.
-
Mosquito Gut Microbiota: A Review.Pathogens. 2024 Aug 15;13(8):691. doi: 10.3390/pathogens13080691. Pathogens. 2024. PMID: 39204291 Free PMC article. Review.
-
An evaluation of fusion partner proteins for paratransgenesis in Asaia bogorensis.PLoS One. 2022 Sep 1;17(9):e0273568. doi: 10.1371/journal.pone.0273568. eCollection 2022. PLoS One. 2022. PMID: 36048823 Free PMC article.
References
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources