

The impact of within-vector parasite development on the extrinsic incubation period
- PMID: 33204441
- PMCID: PMC7657899
- DOI: 10.1098/rsos.192173
The impact of within-vector parasite development on the extrinsic incubation period
Abstract
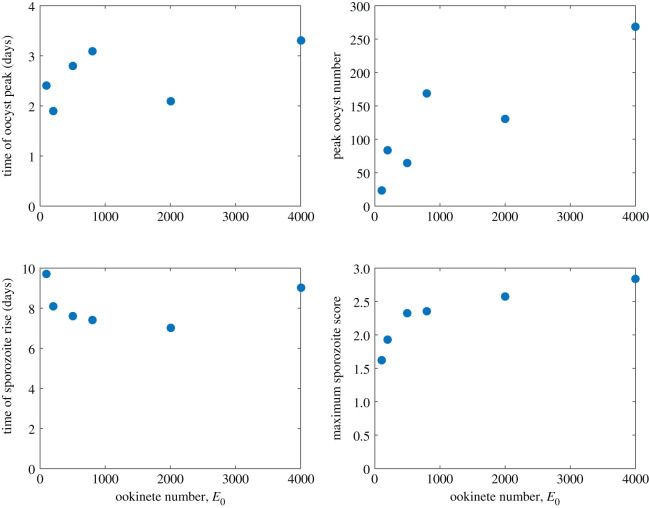
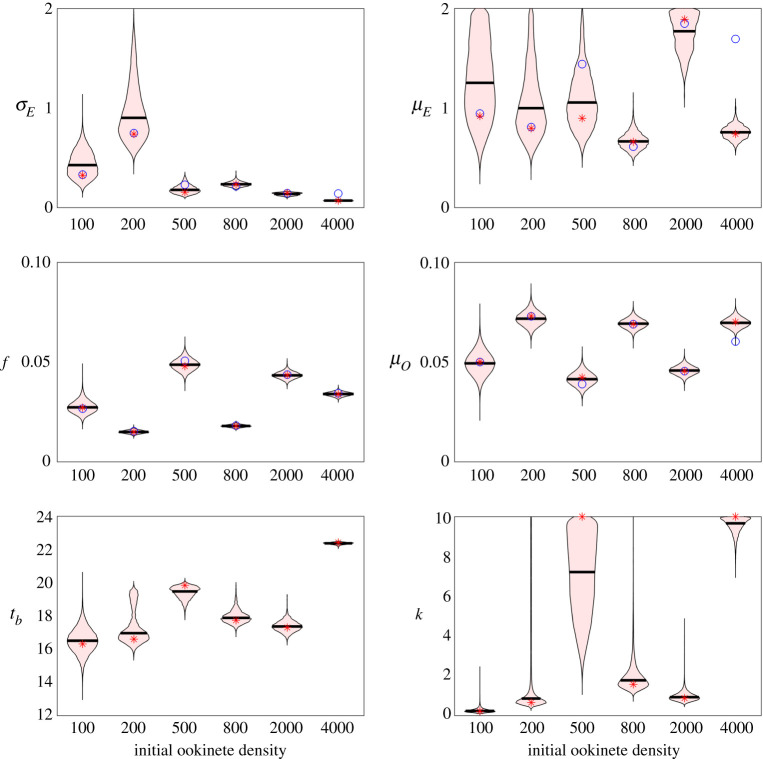
Mosquito-borne diseases, in particular malaria, have a significant burden worldwide leading to nearly half a million deaths each year. The malaria parasite requires a vertebrate host, such as a human, and a vector host, the Anopheles mosquito, to complete its full life cycle. Here, we focus on the parasite dynamics within the vector to examine the first appearance of sporozoites in the salivary glands, which indicates a first time of infectiousness of mosquitoes. The timing of this period of pathogen development in the mosquito until transmissibility, known as the extrinsic incubation period, remains poorly understood. We develop compartmental models of within-mosquito parasite dynamics fitted with experimental data on oocyst and sporozoite counts. We find that only a fraction of oocysts burst to release sporozoites and bursting must be delayed either via a time-dependent function or a gamma-distributed set of compartments. We use Bayesian inference to estimate distributions of parameters and determine that bursting rate is a key epidemiological parameter. A better understanding of the factors impacting the extrinsic incubation period will aid in the development of interventions to slow or stop the spread of malaria.
Keywords: Bayesian inference; extrinsic incubation period; malaria; mosquito; within-host model.
© 2020 The Authors.
Conflict of interest statement
The authors have no competing interests to declare.
Figures





Similar articles
-
Population dynamics of sporogony for Plasmodium vivax parasites from western Thailand developing within three species of colonized Anopheles mosquitoes.Malar J. 2006 Aug 3;5:68. doi: 10.1186/1475-2875-5-68. Malar J. 2006. PMID: 16887043 Free PMC article.
-
Dynamics of Plasmodium vivax sporogony in wild Anopheles stephensi in a malaria-endemic region of Western India.Malar J. 2017 Jul 11;16(1):284. doi: 10.1186/s12936-017-1931-8. Malar J. 2017. PMID: 28693607 Free PMC article.
-
Noninfectious sporozoites in the salivary glands of a minimally susceptible anopheline mosquito.J Parasitol. 1995 Dec;81(6):912-5. J Parasitol. 1995. PMID: 8544063
-
Malaria parasite development in mosquitoes.Annu Rev Entomol. 1998;43:519-43. doi: 10.1146/annurev.ento.43.1.519. Annu Rev Entomol. 1998. PMID: 9444756 Review.
-
Getting infectious: formation and maturation of Plasmodium sporozoites in the Anopheles vector.Cell Microbiol. 2006 Oct;8(10):1547-56. doi: 10.1111/j.1462-5822.2006.00778.x. Cell Microbiol. 2006. PMID: 16984410 Review.
Cited by
-
Investigating the dose-dependency of the midgut escape barrier using a mechanistic model of within-mosquito dengue virus population dynamics.PLoS Pathog. 2024 Apr 1;20(4):e1011975. doi: 10.1371/journal.ppat.1011975. eCollection 2024 Apr. PLoS Pathog. 2024. PMID: 38557892 Free PMC article.
-
The influence of oviposition status on measures of transmission potential in malaria-infected mosquitoes depends on sugar availability.Parasit Vectors. 2024 May 23;17(1):236. doi: 10.1186/s13071-024-06317-2. Parasit Vectors. 2024. PMID: 38783366 Free PMC article.
-
Modeling cellular co-infection and reassortment of bluetongue virus in Culicoides midges.Virus Evol. 2022 Sep 30;8(2):veac094. doi: 10.1093/ve/veac094. eCollection 2022. Virus Evol. 2022. PMID: 36381232 Free PMC article.
-
Omitting age-dependent mosquito mortality in malaria models underestimates the effectiveness of insecticide-treated nets.PLoS Comput Biol. 2022 Sep 19;18(9):e1009540. doi: 10.1371/journal.pcbi.1009540. eCollection 2022 Sep. PLoS Comput Biol. 2022. PMID: 36121847 Free PMC article.
-
Testing a non-destructive assay to track Plasmodium sporozoites in mosquitoes over time.Parasit Vectors. 2023 Nov 4;16(1):401. doi: 10.1186/s13071-023-06015-5. Parasit Vectors. 2023. PMID: 37925480 Free PMC article.
References
-
- WHO. Mosquito-borne diseases: mosquitoes cause millions of deaths every year. https://www.who.int/neglected_diseases/vector_ecology/mosquito-borne-dis....
-
- Teboh-Ewungkem MI, Yuster T, Newman NH. 2010. A mathematical model of the within-vector dynamics of the Plasmodium falciparum protozoan parasite. In Infectious disease modelling research progress (eds JM Tchuenche, C Chiyaka), pp. 177–199. New York, NY: Nova Science Publishers.
Associated data
LinkOut - more resources
Full Text Sources
