

Mitochondrial metabolic substrate utilization in granulosa cells reflects body mass index and total follicle stimulating hormone dosage in in vitro fertilization patients
- PMID: 32935173
- PMCID: PMC7642160
- DOI: 10.1007/s10815-020-01946-9
Mitochondrial metabolic substrate utilization in granulosa cells reflects body mass index and total follicle stimulating hormone dosage in in vitro fertilization patients
Abstract
Purpose: To utilize a novel mitochondrial function assay with pooled granulosa cells to determine whether mitochondrial function would differ by patient demographics and embryo development.
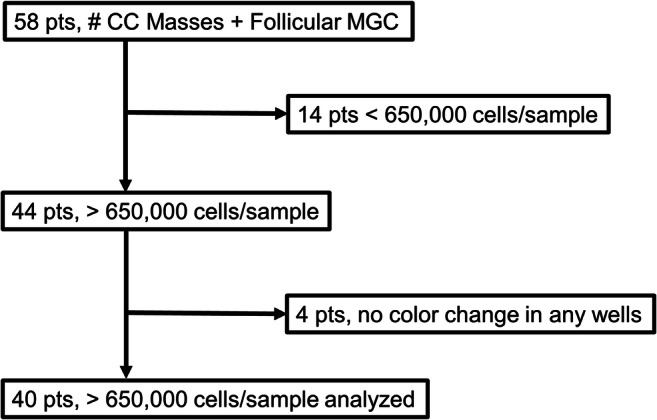
Methods: This was a prospective pilot study in a hospital-based assisted reproductive program and public university. Mitochondrial metabolic substrate utilization was assessed in pooled granulosa cells from 40 women undergoing in vitro fertilization during 2018 and 2019.
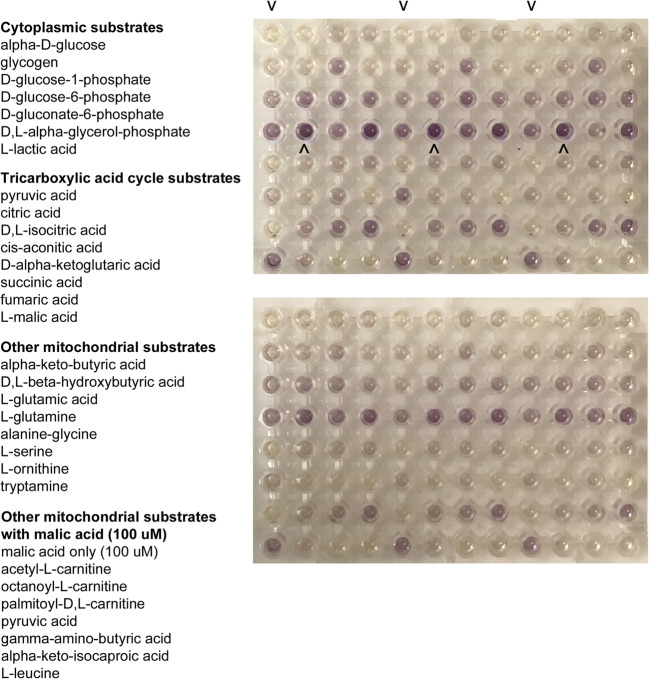
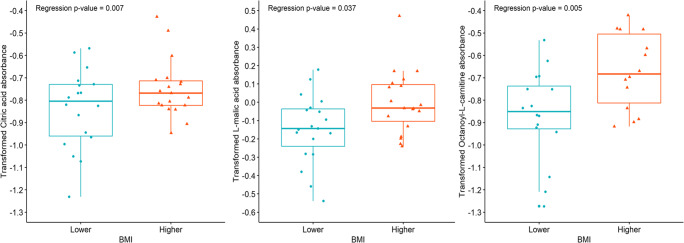
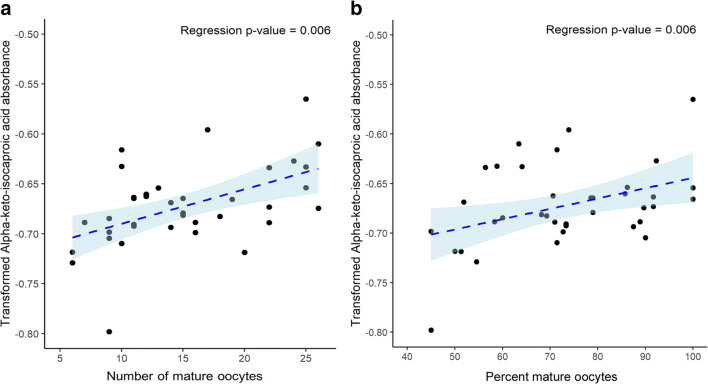
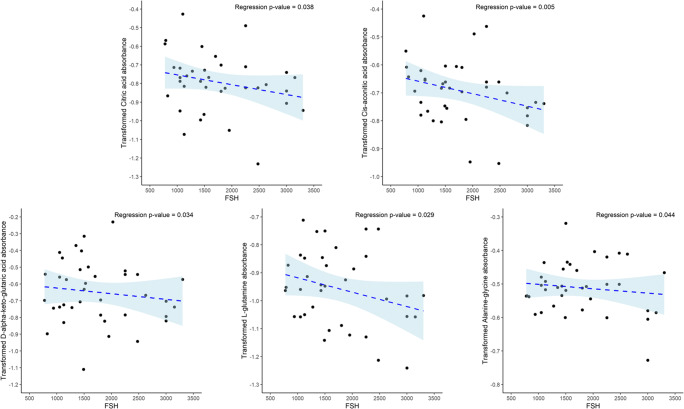
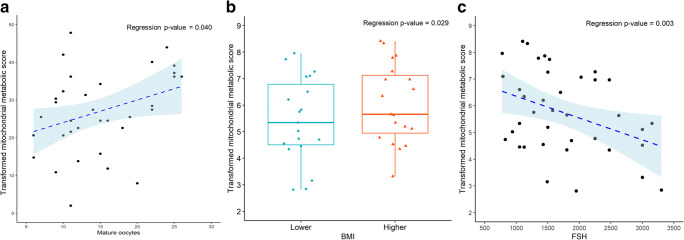
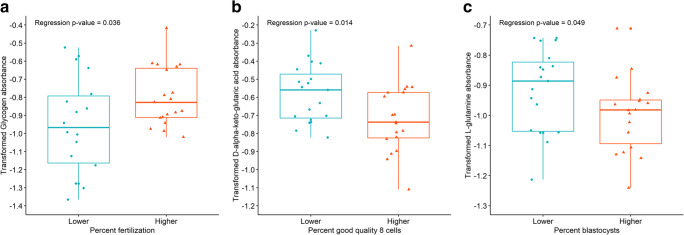
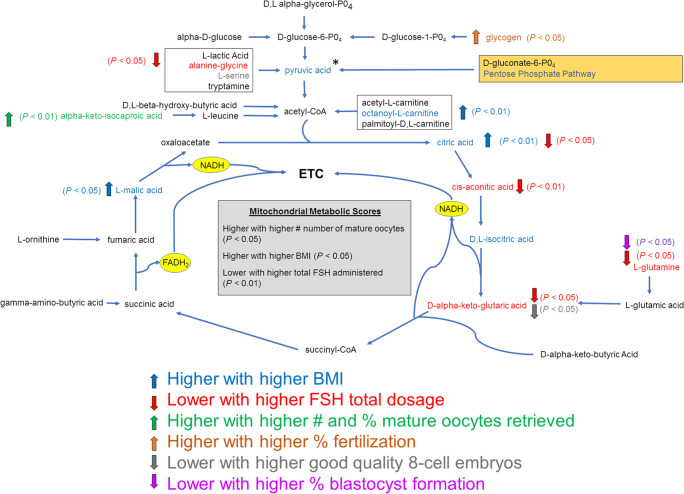
Results: Assessment of mitochondrial substrate metabolism in pooled granulosa cells revealed higher citric acid, L-malic acid, and octanoyl-L-carnitine utilization with higher body mass index (BMI). Utilization of citric acid, cis-aconitic acid, D-alpha-keto-glutaric acid, L-glutamine, and alanine plus glycine was significantly lower as total dosage of FSH administered increased. Utilization of glycogen was significantly higher in patients with a higher percentage of fertilized oocytes. D-alpha-keto-glutaric acid utilization was significantly lower in patients with a higher percentage of good 8-cell embryos. L-glutamine utilization was significantly lower, with a higher percentage of blastocyst formation. Mitochondrial metabolic scores (MMS), which reflect overall mitochondrial activity of the granulosa pool, were significantly higher in patients with higher BMI and with greater numbers of mature oocytes retrieved. MMS in granulosa decreased as total FSH dose administered increased.
Conclusions: Granulosa cell utilization of substrates feeding into the citric acid cycle changed with total FSH dosage and BMI. Fertilization rate, 8-cell embryo quality, and blastocyst formation also associated with different energy substrate usage. Mitochondrial substrate utilization by granulosa cells from individual follicles could be further developed into a useful diagnostic tool.
Keywords: Citric acid cycle; Granulosa cells; In vitro fertilization; Mitochondrial function assay; Mitochondrial metabolic score; Mitochondrial substrate utilization.
Conflict of interest statement
The authors declare no conflict of interest related to this study.
Figures
Similar articles
-
Mitochondrial Function in Modulating Human Granulosa Cell Steroidogenesis and Female Fertility.Int J Mol Sci. 2020 May 19;21(10):3592. doi: 10.3390/ijms21103592. Int J Mol Sci. 2020. PMID: 32438750 Free PMC article.
-
Oocyte matched follicular fluid anti-Müllerian hormone is an excellent predictor of live birth after fresh single embryo transfer.Hum Reprod. 2019 Nov 1;34(11):2244-2253. doi: 10.1093/humrep/dez186. Hum Reprod. 2019. PMID: 31725884
-
A pre-in vitro maturation medium containing cumulus oocyte complex ligand-receptor signaling molecules maintains meiotic arrest, supports the cumulus oocyte complex and improves oocyte developmental competence.Mol Hum Reprod. 2017 Sep 1;23(9):594-606. doi: 10.1093/molehr/gax032. Mol Hum Reprod. 2017. PMID: 28586460
-
Ovarian follicular and luteal physiology.Int Rev Physiol. 1980;22:117-201. Int Rev Physiol. 1980. PMID: 6248477 Review.
-
Mechanism of Mitochondrial Homeostasis Controlling Ovarian Physiology.Endocrinology. 2022 Nov 14;164(1):bqac189. doi: 10.1210/endocr/bqac189. Endocrinology. 2022. PMID: 36378567 Review.
Cited by
-
High body energy reserve influences extracellular vesicles miRNA contents within the ovarian follicle.PLoS One. 2023 Jan 10;18(1):e0280195. doi: 10.1371/journal.pone.0280195. eCollection 2023. PLoS One. 2023. PMID: 36626404 Free PMC article.
-
Omics data integration suggests a potential idiopathic Parkinson's disease signature.Commun Biol. 2023 Nov 20;6(1):1179. doi: 10.1038/s42003-023-05548-w. Commun Biol. 2023. PMID: 37985891 Free PMC article.
-
Modelling Metabolic Shifts during Cardiomyocyte Differentiation, Iron Deficiency and Transferrin Rescue Using Human Pluripotent Stem Cells.Metabolites. 2021 Dec 22;12(1):9. doi: 10.3390/metabo12010009. Metabolites. 2021. PMID: 35050131 Free PMC article.
-
Granulosa cell metabolism at ovulation correlates with oocyte competence and is disrupted by obesity and aging.Hum Reprod. 2024 Sep 1;39(9):2053-2066. doi: 10.1093/humrep/deae154. Hum Reprod. 2024. PMID: 39013118 Free PMC article.
-
Maturational competence of equine oocytes is associated with alterations in their 'cumulome'.Mol Hum Reprod. 2024 Sep 12;30(9):gaae033. doi: 10.1093/molehr/gaae033. Mol Hum Reprod. 2024. PMID: 39288330 Free PMC article.
References
-
- Boucret L, JM CdlB, Moriniere C, Desquiret V, Ferre-L'Hotellier V, Descamps P et al. Relationship between diminished ovarian reserve and mitochondrial biogenesis in cumulus cells. Hum Reprod. 2015;30(7):1653–64. doi:dev114 [pii];10.1093/humrep/dev114. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
