

Gut microbiota dysbiosis and altered tryptophan catabolism contribute to autoimmunity in lupus-susceptible mice
- PMID: 32641487
- PMCID: PMC7739186
- DOI: 10.1126/scitranslmed.aax2220
Gut microbiota dysbiosis and altered tryptophan catabolism contribute to autoimmunity in lupus-susceptible mice
Abstract
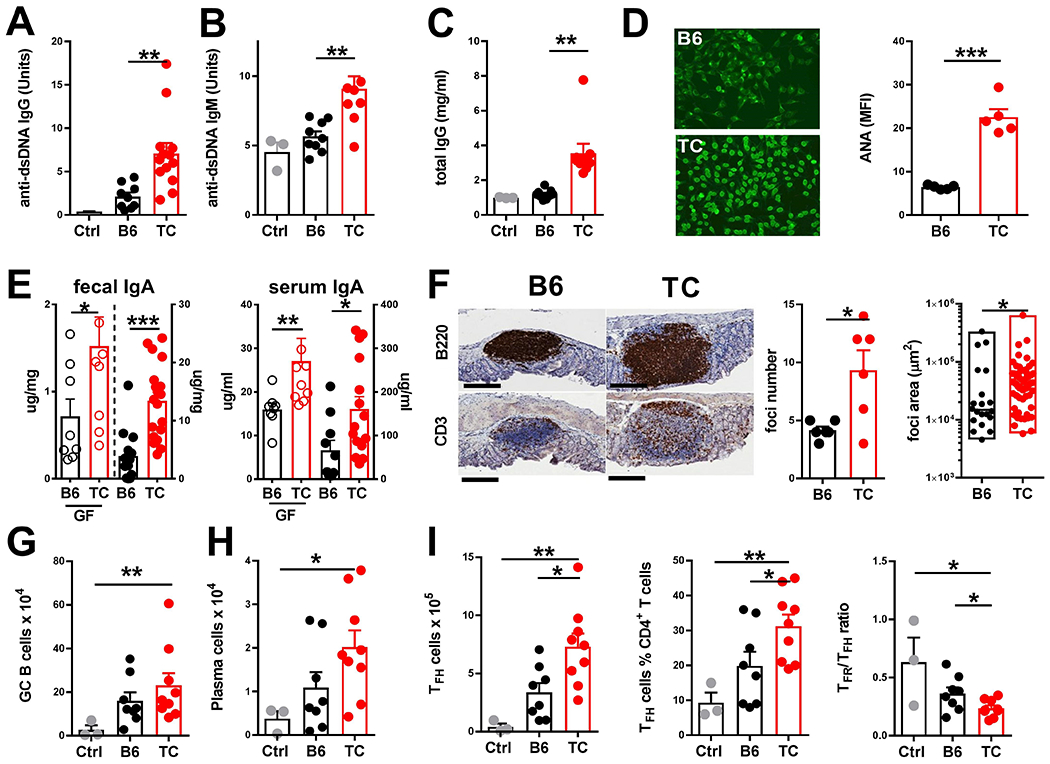
The autoimmune disease systemic lupus erythematosus (SLE) is characterized by the production of pathogenic autoantibodies. It has been postulated that gut microbial dysbiosis may be one of the mechanisms involved in SLE pathogenesis. Here, we demonstrate that the dysbiotic gut microbiota of triple congenic (TC) lupus-prone mice (B6.Sle1.Sle2.Sle3) stimulated the production of autoantibodies and activated immune cells when transferred into germfree congenic C57BL/6 (B6) mice. Fecal transfer to B6 mice induced autoimmune phenotypes only when the TC donor mice exhibited autoimmunity. Autoimmune pathogenesis was mitigated by horizontal transfer of the gut microbiota between co-housed lupus-prone TC mice and control congenic B6 mice. Metabolomic screening identified an altered distribution of tryptophan metabolites in the feces of TC mice including an increase in kynurenine, which was alleviated after antibiotic treatment. Low dietary tryptophan prevented autoimmune pathology in TC mice, whereas high dietary tryptophan exacerbated disease. Reducing dietary tryptophan altered gut microbial taxa in both lupus-prone TC mice and control B6 mice. Consequently, fecal transfer from TC mice fed a high tryptophan diet, but not a low tryptophan diet, induced autoimmune phenotypes in germfree B6 mice. The interplay of gut microbial dysbiosis, tryptophan metabolism and host genetic susceptibility in lupus-prone mice suggest that aberrant tryptophan metabolism may contribute to autoimmune activation in this disease.
Copyright © 2020 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Conflict of interest statement
Figures





Comment in
-
Microbiota, metabolism and lupus in mice.Nat Rev Rheumatol. 2020 Sep;16(9):474. doi: 10.1038/s41584-020-0482-5. Nat Rev Rheumatol. 2020. PMID: 32704078 No abstract available.
Similar articles
-
Dietary tryptophan and genetic susceptibility expand gut microbiota that promote systemic autoimmune activation.bioRxiv [Preprint]. 2024 Jan 19:2024.01.16.575942. doi: 10.1101/2024.01.16.575942. bioRxiv. 2024. PMID: 38293097 Free PMC article. Preprint.
-
Bidirectional crosstalk between dysbiotic gut microbiota and systemic lupus erythematosus: What is new in therapeutic approaches?Clin Immunol. 2022 Nov;244:109109. doi: 10.1016/j.clim.2022.109109. Epub 2022 Sep 8. Clin Immunol. 2022. PMID: 36087683 Review.
-
Genetic reconstitution of systemic lupus erythematosus immunopathology with polycongenic murine strains.Proc Natl Acad Sci U S A. 2000 Jun 6;97(12):6670-5. doi: 10.1073/pnas.97.12.6670. Proc Natl Acad Sci U S A. 2000. PMID: 10841565 Free PMC article.
-
Intestinal Dysbiosis and Tryptophan Metabolism in Autoimmunity.Front Immunol. 2020 Aug 4;11:1741. doi: 10.3389/fimmu.2020.01741. eCollection 2020. Front Immunol. 2020. PMID: 32849620 Free PMC article. Review.
-
Lupus gut microbiota transplants cause autoimmunity and inflammation.Clin Immunol. 2021 Dec;233:108892. doi: 10.1016/j.clim.2021.108892. Epub 2021 Nov 20. Clin Immunol. 2021. PMID: 34813937
Cited by
-
Tryptophan Metabolism and Gut-Brain Homeostasis.Int J Mol Sci. 2021 Mar 15;22(6):2973. doi: 10.3390/ijms22062973. Int J Mol Sci. 2021. PMID: 33804088 Free PMC article. Review.
-
Immunometabolic alterations in lupus: where do they come from and where do we go from there?Curr Opin Immunol. 2022 Oct;78:102245. doi: 10.1016/j.coi.2022.102245. Epub 2022 Sep 16. Curr Opin Immunol. 2022. PMID: 36122544 Free PMC article. Review.
-
Vitamin B12 Regulates the Transcriptional, Metabolic, and Epigenetic Programing in Human Ileal Epithelial Cells.Nutrients. 2022 Jul 9;14(14):2825. doi: 10.3390/nu14142825. Nutrients. 2022. PMID: 35889782 Free PMC article.
-
A Scoping Review of the Positive and Negative Bacteria Associated With the Gut Microbiomes of Systemic Lupus Erythematosus Patients.Cureus. 2024 Apr 3;16(4):e57512. doi: 10.7759/cureus.57512. eCollection 2024 Apr. Cureus. 2024. PMID: 38707123 Free PMC article.
-
Causal Effects of Gut Microbiome on Systemic Lupus Erythematosus: A Two-Sample Mendelian Randomization Study.Front Immunol. 2021 Sep 7;12:667097. doi: 10.3389/fimmu.2021.667097. eCollection 2021. Front Immunol. 2021. PMID: 34557183 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
