

Spatial and temporal alterations in protein structure by EGF regulate cryptic cysteine oxidation
- PMID: 31964804
- PMCID: PMC7263378
- DOI: 10.1126/scisignal.aay7315
Spatial and temporal alterations in protein structure by EGF regulate cryptic cysteine oxidation
Abstract
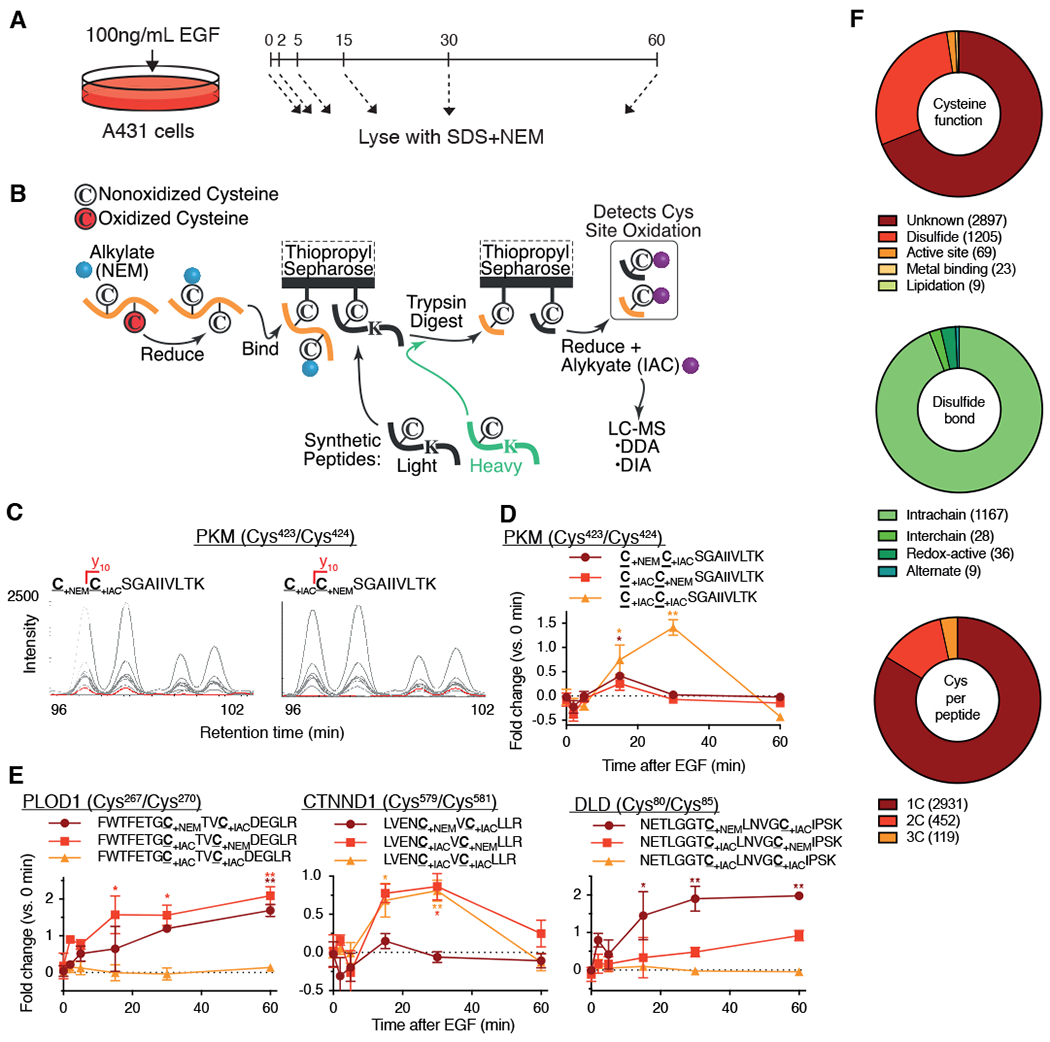
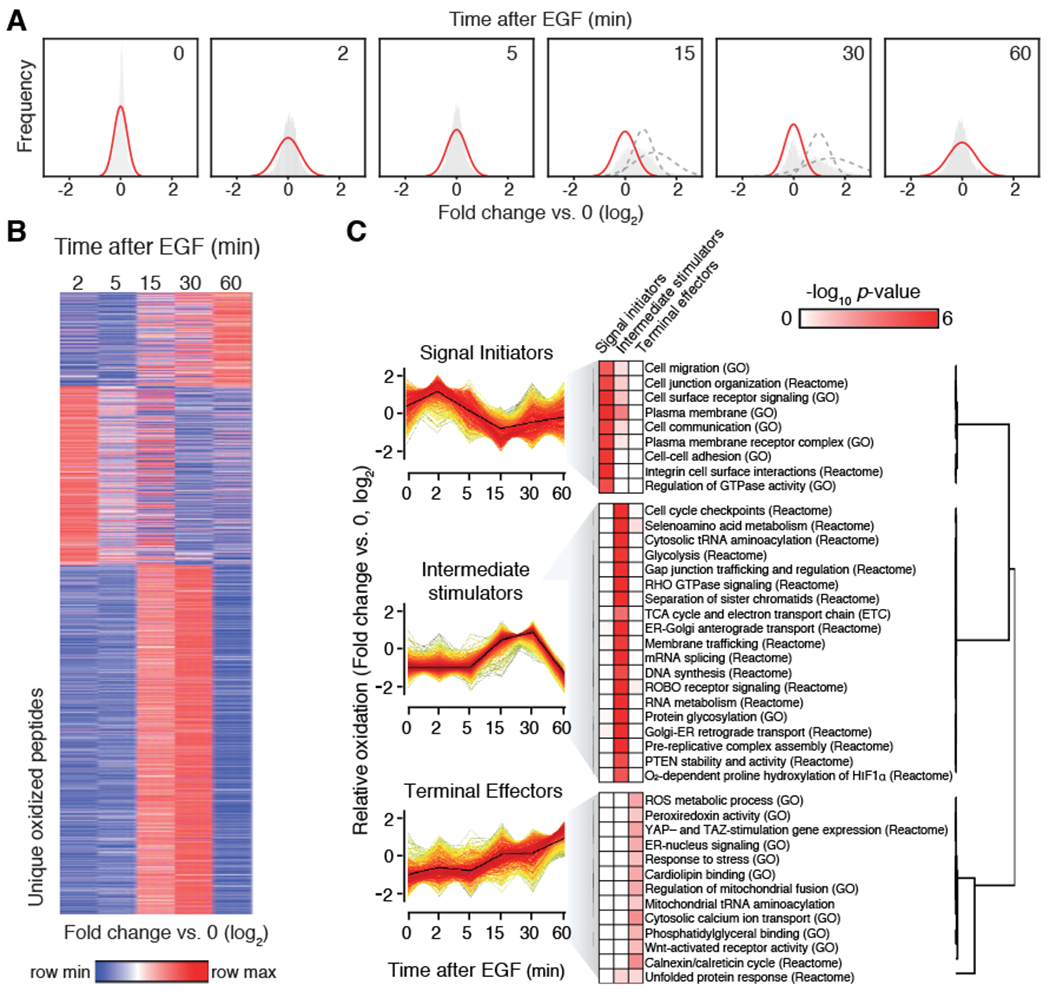
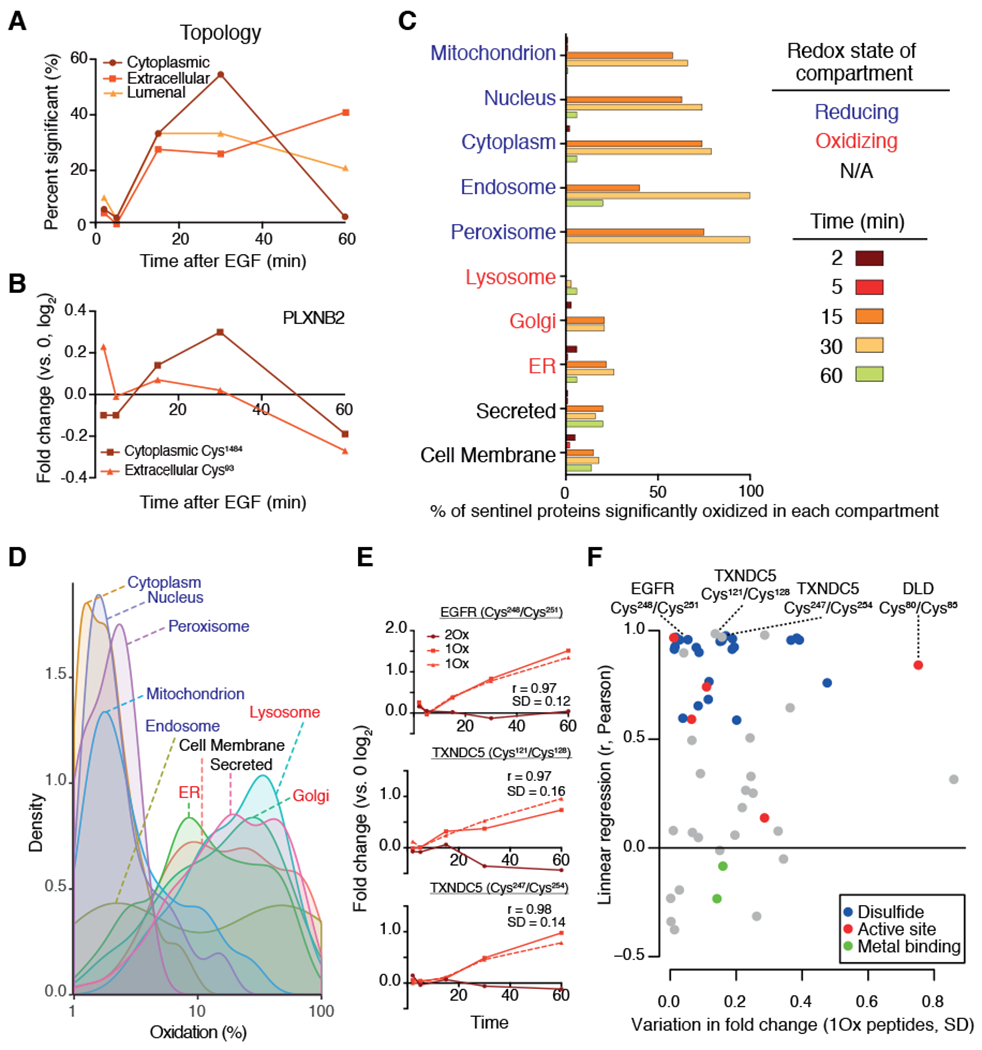
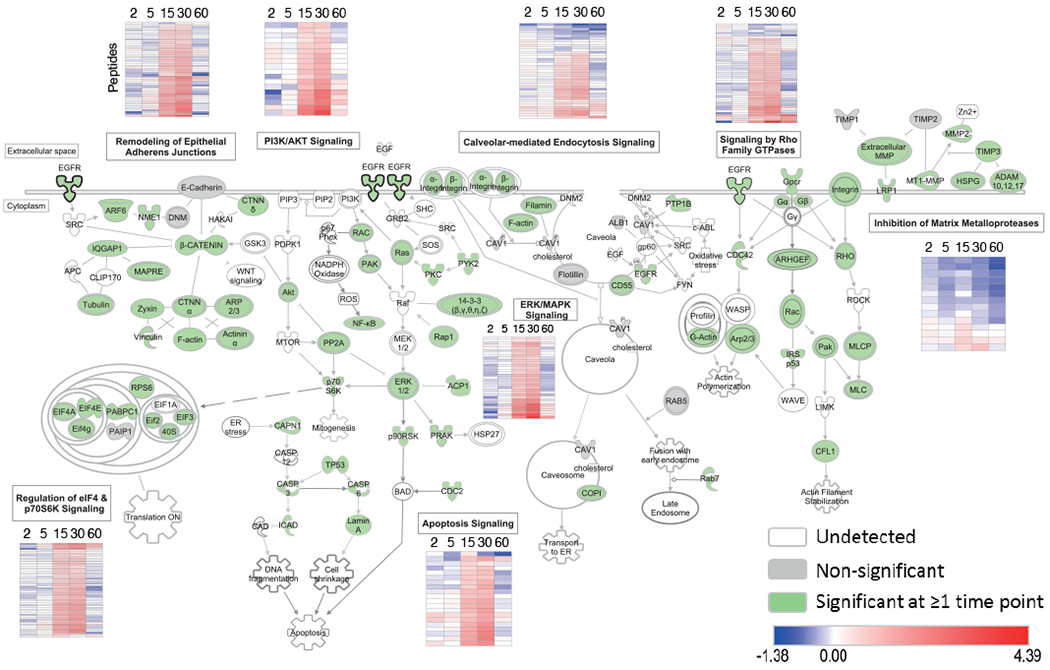
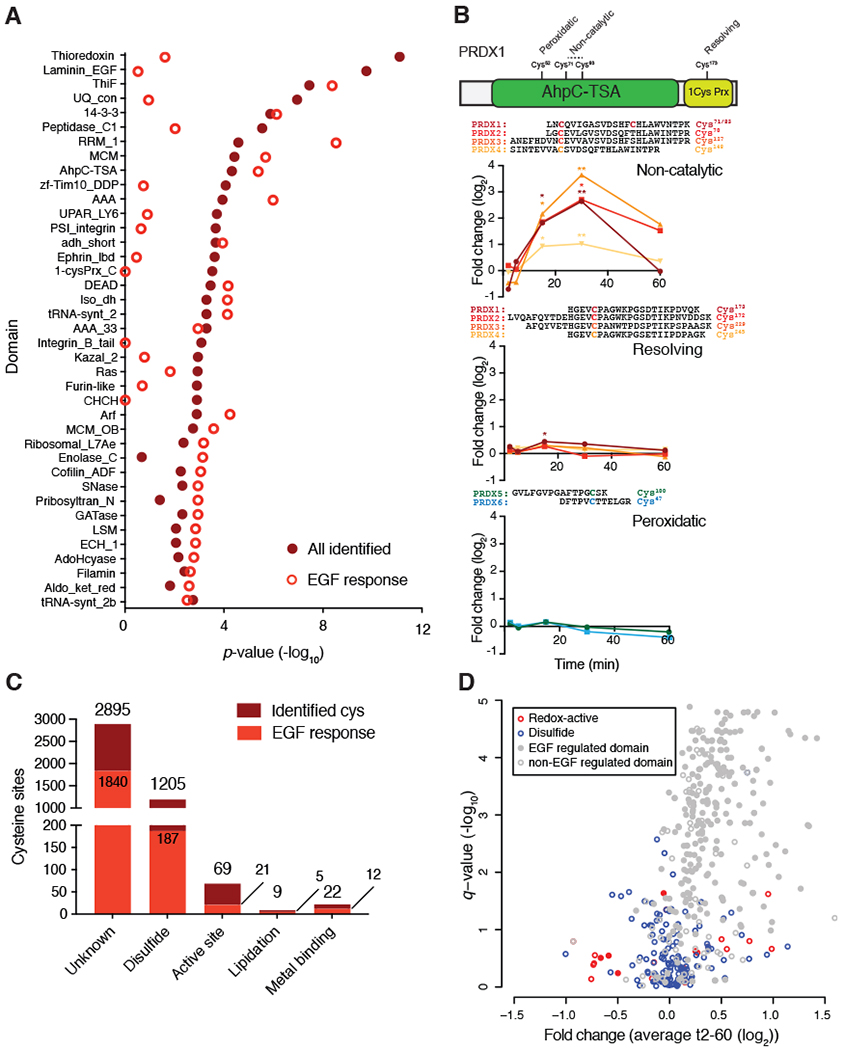
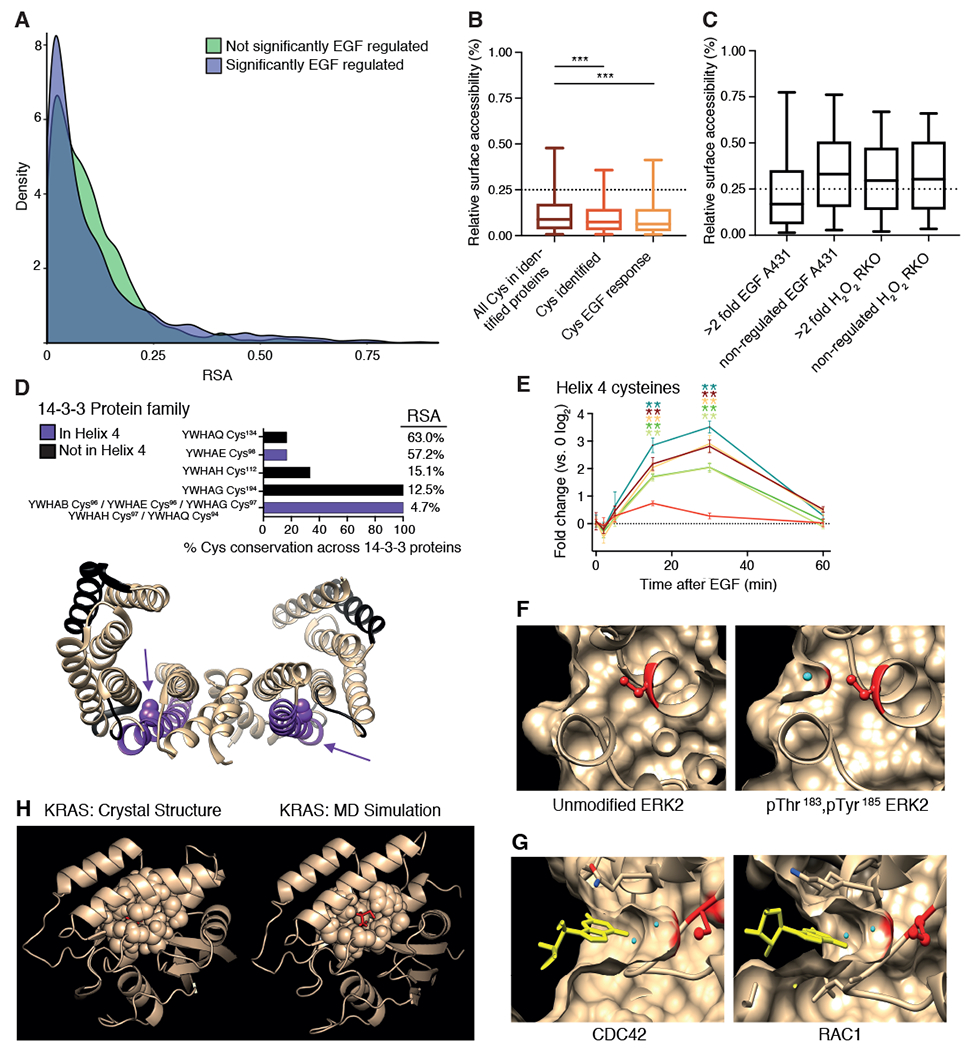
Stimulation of plasma membrane receptor tyrosine kinases (RTKs), such as the epidermal growth factor receptor (EGFR), locally increases the abundance of reactive oxygen species (ROS). These ROS then oxidize cysteine residues in proteins to potentiate downstream signaling. Spatial confinement of ROS is an important regulatory mechanism of redox signaling that enables the stimulation of different RTKs to oxidize distinct sets of downstream proteins. To uncover additional mechanisms that specify cysteines that are redox regulated by EGF stimulation, we performed time-resolved quantification of the EGF-dependent oxidation of 4200 cysteine sites in A431 cells. Fifty-one percent of cysteines were statistically significantly oxidized by EGF stimulation. Furthermore, EGF induced three distinct spatiotemporal patterns of cysteine oxidation in functionally organized protein networks, consistent with the spatial confinement model. Unexpectedly, protein crystal structure analysis and molecular dynamics simulations indicated widespread redox regulation of cryptic cysteine residues that are solvent exposed only upon changes in protein conformation. Phosphorylation and increased flux of nucleotide substrates served as two distinct modes by which EGF specified the cryptic cysteine residues that became solvent exposed and redox regulated. Because proteins that are structurally regulated by different RTKs or cellular perturbations are largely unique, these findings suggest that solvent exposure and redox regulation of cryptic cysteine residues contextually delineate redox signaling networks.
Copyright © 2020 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Conflict of interest statement
Figures
Similar articles
-
Differential activation of epidermal growth factor (EGF) receptor downstream signaling pathways by betacellulin and EGF.Endocrinology. 2004 Sep;145(9):4232-43. doi: 10.1210/en.2004-0401. Epub 2004 Jun 10. Endocrinology. 2004. PMID: 15192046
-
The NADPH Oxidases DUOX1 and NOX2 Play Distinct Roles in Redox Regulation of Epidermal Growth Factor Receptor Signaling.J Biol Chem. 2016 Oct 28;291(44):23282-23293. doi: 10.1074/jbc.M116.749028. Epub 2016 Sep 20. J Biol Chem. 2016. PMID: 27650496 Free PMC article.
-
Steric constraints in the recognition of peptide substrates for the epidermal growth factor receptor kinase.Int J Pept Protein Res. 1996 Mar;47(3):219-26. doi: 10.1111/j.1399-3011.1996.tb01348.x. Int J Pept Protein Res. 1996. PMID: 8740973
-
Redox regulation of epidermal growth factor receptor signaling through cysteine oxidation.Biochemistry. 2012 Dec 18;51(50):9954-65. doi: 10.1021/bi301441e. Epub 2012 Dec 5. Biochemistry. 2012. PMID: 23186290 Free PMC article. Review.
-
Redox-dependent regulation of epidermal growth factor receptor signaling.Redox Biol. 2016 Aug;8:24-7. doi: 10.1016/j.redox.2015.12.002. Epub 2015 Dec 8. Redox Biol. 2016. PMID: 26722841 Free PMC article. Review.
Cited by
-
A clickable probe for versatile characterization of S-nitrosothiols.Redox Biol. 2020 Oct;37:101707. doi: 10.1016/j.redox.2020.101707. Epub 2020 Sep 1. Redox Biol. 2020. PMID: 32916549 Free PMC article.
-
Automating Assignment, Quantitation, and Biological Annotation of Redox Proteomics Datasets with ProteoSushi.Methods Mol Biol. 2022;2399:61-84. doi: 10.1007/978-1-0716-1831-8_4. Methods Mol Biol. 2022. PMID: 35604553
-
Proteins in Tumor-Derived Plasma Extracellular Vesicles Indicate Tumor Origin.Mol Cell Proteomics. 2023 Jan;22(1):100476. doi: 10.1016/j.mcpro.2022.100476. Epub 2022 Dec 5. Mol Cell Proteomics. 2023. PMID: 36470535 Free PMC article.
-
NOX1-dependent redox signaling potentiates colonic stem cell proliferation to adapt to the intestinal microbiota by linking EGFR and TLR activation.Cell Rep. 2021 Apr 6;35(1):108949. doi: 10.1016/j.celrep.2021.108949. Cell Rep. 2021. PMID: 33826887 Free PMC article.
-
Characterization of cellular oxidative stress response by stoichiometric redox proteomics.Am J Physiol Cell Physiol. 2021 Feb 1;320(2):C182-C194. doi: 10.1152/ajpcell.00040.2020. Epub 2020 Dec 2. Am J Physiol Cell Physiol. 2021. PMID: 33264075 Free PMC article. Review.
References
-
- Bae YS, Kang SW, Seo MS, Baines IC, Tekle E, Chock PB, Rhee SG, Epidermal Growth Factor (EGF)-induced Generation of Hydrogen Peroxide, J. Biol. Chem 272, 217–221 (1997). - PubMed
-
- Winterbourn CC, Reconciling the chemistry and biology of reactive oxygen species, Nat. Chem. Biol 4, 278–286 (2008). - PubMed
-
- Mahadev K, Zilbering A, Zhu L, Goldstein BJ, Insulin-stimulated Hydrogen Peroxide Reversibly Inhibits Protein-tyrosine Phosphatase 1B in Vivo and Enhances the Early Insulin Action Cascade, J. Biol. Chem 276, 21938–21942 (2001). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
