

Bridging the gap between single-cell migration and collective dynamics
- PMID: 31808744
- PMCID: PMC6992385
- DOI: 10.7554/eLife.46842
Bridging the gap between single-cell migration and collective dynamics
Abstract
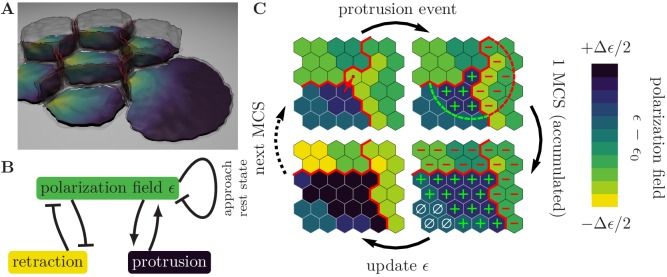
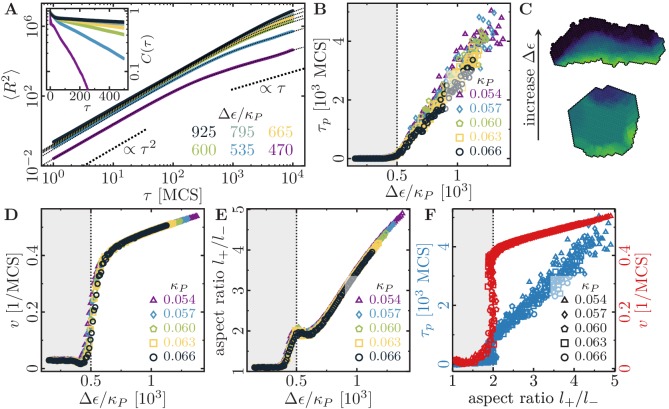
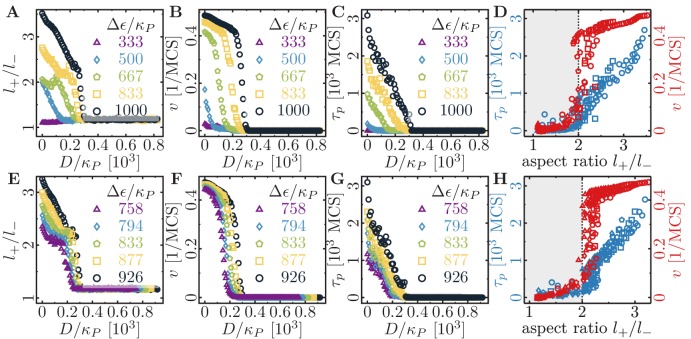
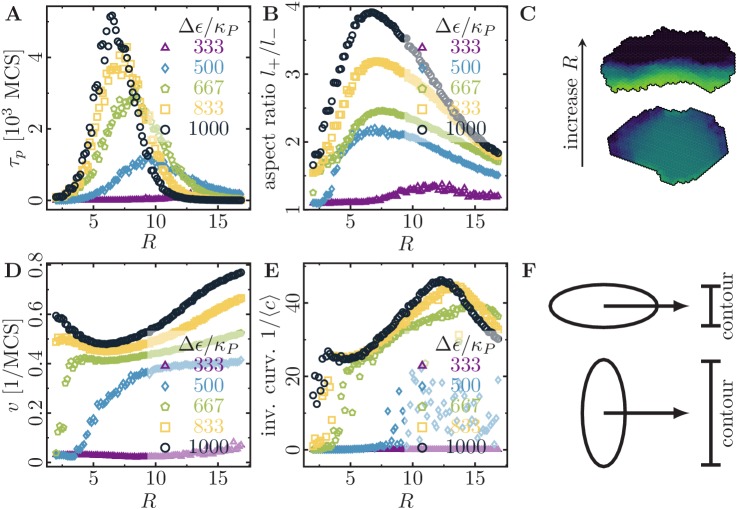
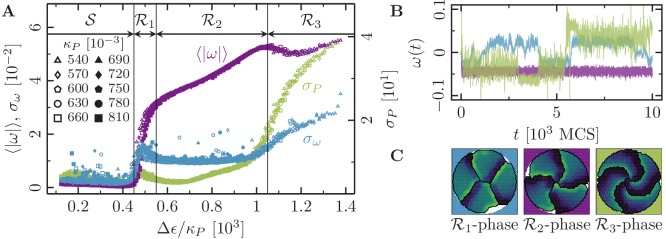
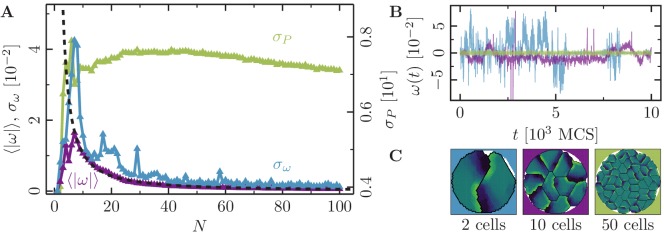
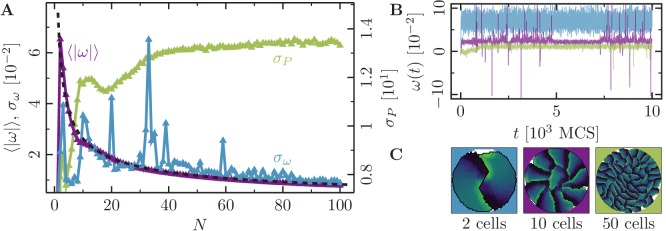
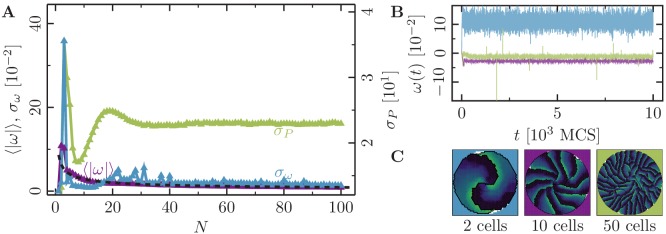
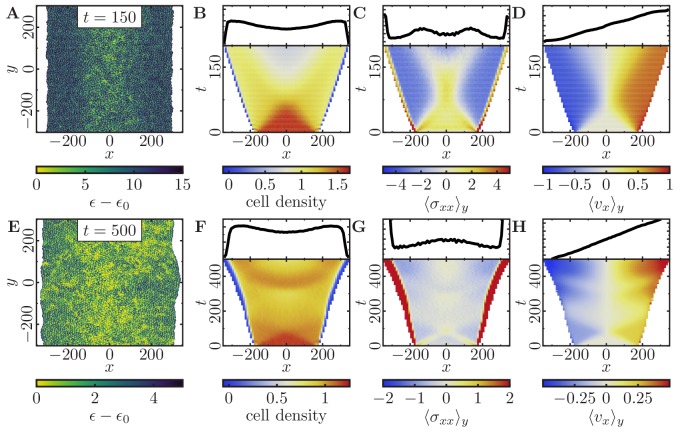
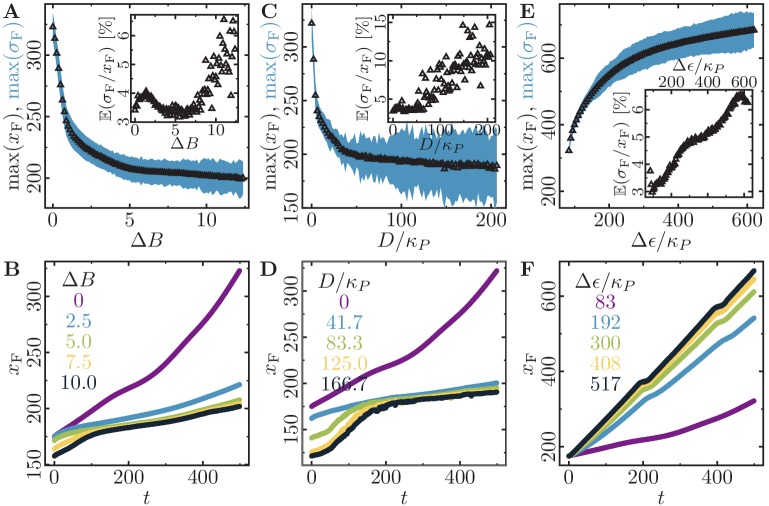
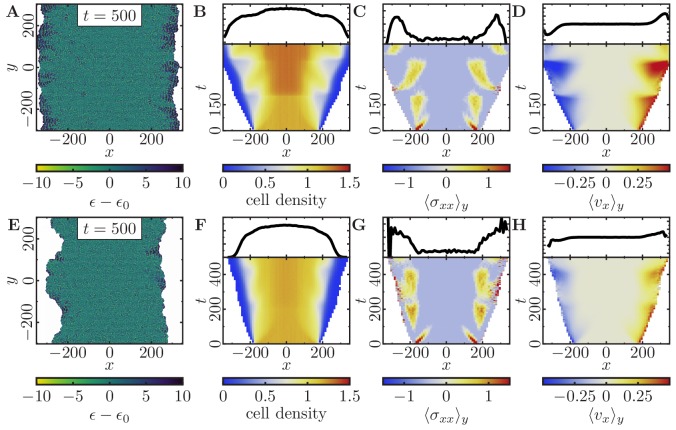
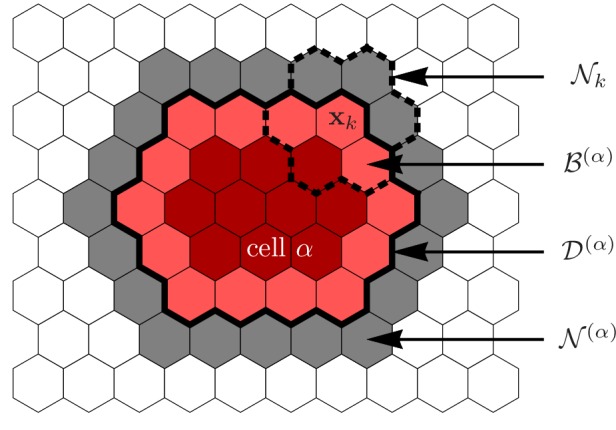
Motivated by the wealth of experimental data recently available, we present a cellular-automaton-based modeling framework focussing on high-level cell functions and their concerted effect on cellular migration patterns. Specifically, we formulate a coarse-grained description of cell polarity through self-regulated actin organization and its response to mechanical cues. Furthermore, we address the impact of cell adhesion on collective migration in cell cohorts. The model faithfully reproduces typical cell shapes and movements down to the level of single cells, yet allows for the efficient simulation of confluent tissues. In confined circular geometries, we find that specific properties of individual cells (polarizability; contractility) influence the emerging collective motion of small cell cohorts. Finally, we study the properties of expanding cellular monolayers (front morphology; stress and velocity distributions) at the level of extended tissues.
Keywords: biophysics; cell biology; cell migration; computational biology; none; physics of living systems; tissue mechanics.
© 2019, Thüroff et al.
Conflict of interest statement
FT, AG, MR, EF No competing interests declared
Figures
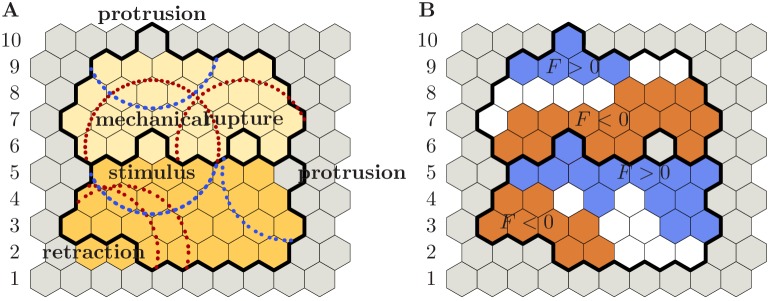
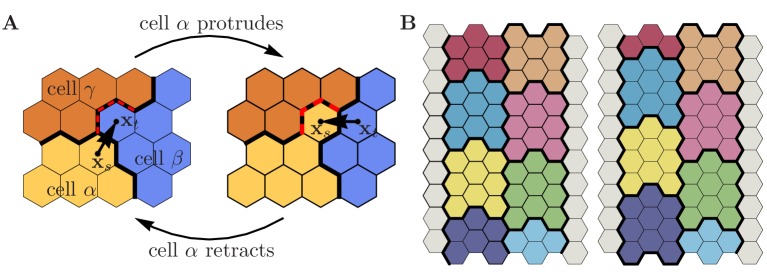
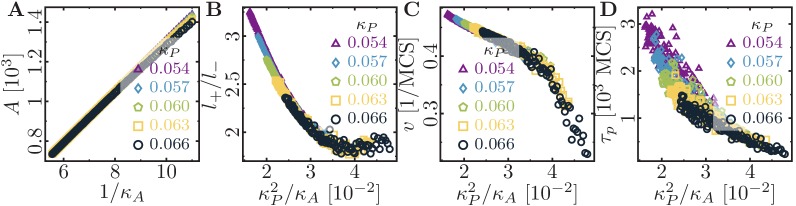
Similar articles
-
Dynamics of Cell Ensembles on Adhesive Micropatterns: Bridging the Gap between Single Cell Spreading and Collective Cell Migration.PLoS Comput Biol. 2016 Apr 7;12(4):e1004863. doi: 10.1371/journal.pcbi.1004863. eCollection 2016 Apr. PLoS Comput Biol. 2016. PMID: 27054883 Free PMC article.
-
Active Vertex Model for cell-resolution description of epithelial tissue mechanics.PLoS Comput Biol. 2017 Jun 30;13(6):e1005569. doi: 10.1371/journal.pcbi.1005569. eCollection 2017 Jun. PLoS Comput Biol. 2017. PMID: 28665934 Free PMC article.
-
Force localization modes in dynamic epithelial colonies.Mol Biol Cell. 2018 Nov 15;29(23):2835-2847. doi: 10.1091/mbc.E18-05-0336. Epub 2018 Sep 12. Mol Biol Cell. 2018. PMID: 30207837 Free PMC article.
-
Tissue patterning and cellular mechanics.J Cell Biol. 2015 Oct 26;211(2):219-31. doi: 10.1083/jcb.201506106. J Cell Biol. 2015. PMID: 26504164 Free PMC article. Review.
-
Phenomenological approaches to collective behavior in epithelial cell migration.Biochim Biophys Acta. 2015 Nov;1853(11 Pt B):3143-52. doi: 10.1016/j.bbamcr.2015.05.021. Epub 2015 May 29. Biochim Biophys Acta. 2015. PMID: 26028592 Review.
Cited by
-
Synthetic Cell as a Platform for Understanding Membrane-Membrane Interactions.Membranes (Basel). 2021 Nov 23;11(12):912. doi: 10.3390/membranes11120912. Membranes (Basel). 2021. PMID: 34940413 Free PMC article. Review.
-
Plasticity of cancer cell invasion: Patterns and mechanisms.Transl Oncol. 2021 Jan;14(1):100899. doi: 10.1016/j.tranon.2020.100899. Epub 2020 Oct 17. Transl Oncol. 2021. PMID: 33080522 Free PMC article. Review.
-
Paxillin promotes breast tumor collective cell invasion through maintenance of adherens junction integrity.Mol Biol Cell. 2022 Feb 1;33(2):ar14. doi: 10.1091/mbc.E21-09-0432. Epub 2021 Dec 1. Mol Biol Cell. 2022. PMID: 34851720 Free PMC article.
-
Multiphoton excited polymerized biomimetic models of collagen fiber morphology to study single cell and collective migration dynamics in pancreatic cancer.Acta Biomater. 2024 Oct 1;187:212-226. doi: 10.1016/j.actbio.2024.08.026. Epub 2024 Aug 23. Acta Biomater. 2024. PMID: 39182805
-
Unravelling cell migration: defining movement from the cell surface.Cell Adh Migr. 2022 Dec;16(1):25-64. doi: 10.1080/19336918.2022.2055520. Cell Adh Migr. 2022. PMID: 35499121 Free PMC article. Review.
References
-
- Alberts B, Johnson A, Lewis J, Raff M. Molecular Biology of the Cell. Sixth Edition . Garland Science; 2015.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
