

Nucleo-cytoplasmic Partitioning of ARF Proteins Controls Auxin Responses in Arabidopsis thaliana
- PMID: 31421981
- PMCID: PMC6778021
- DOI: 10.1016/j.molcel.2019.06.044
Nucleo-cytoplasmic Partitioning of ARF Proteins Controls Auxin Responses in Arabidopsis thaliana
Abstract
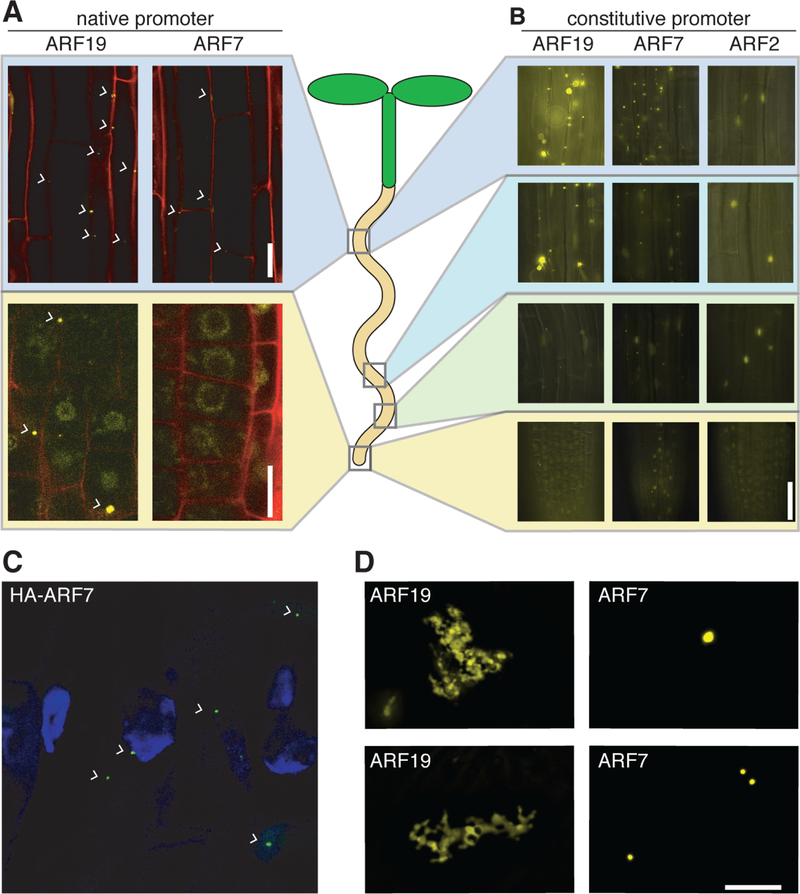
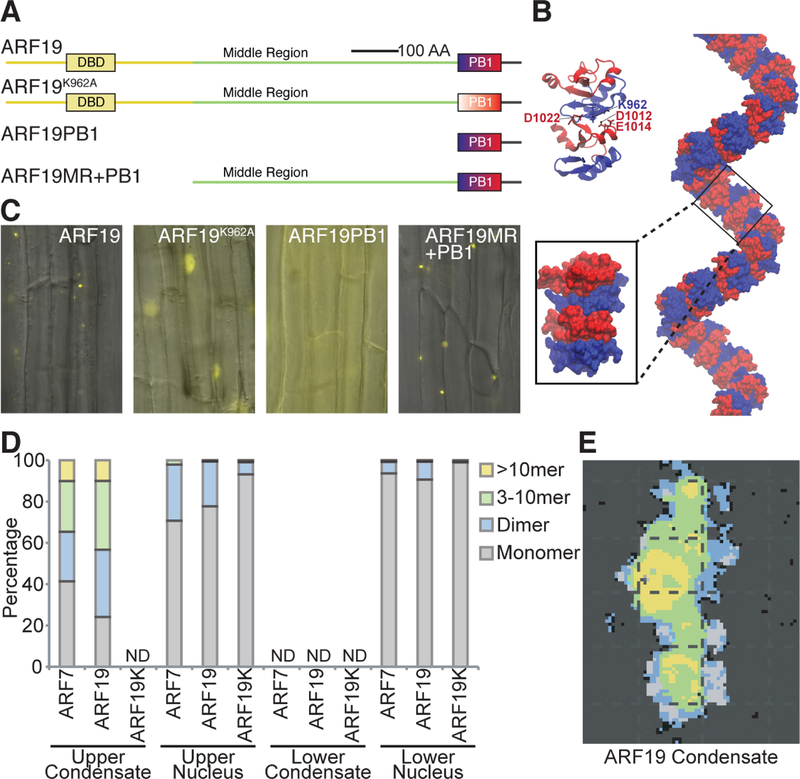
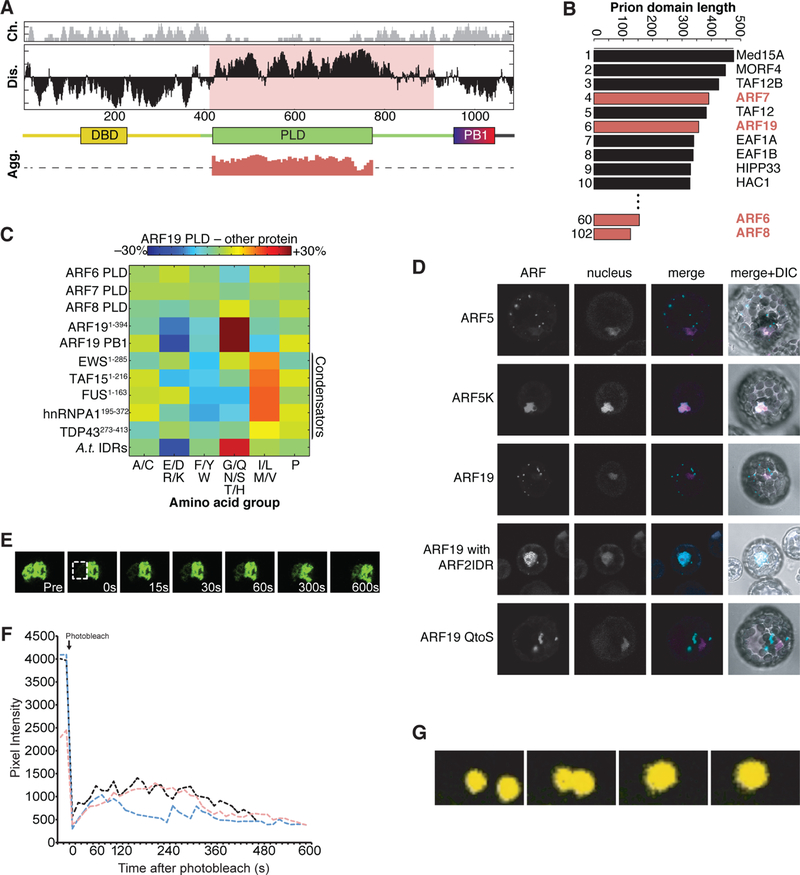
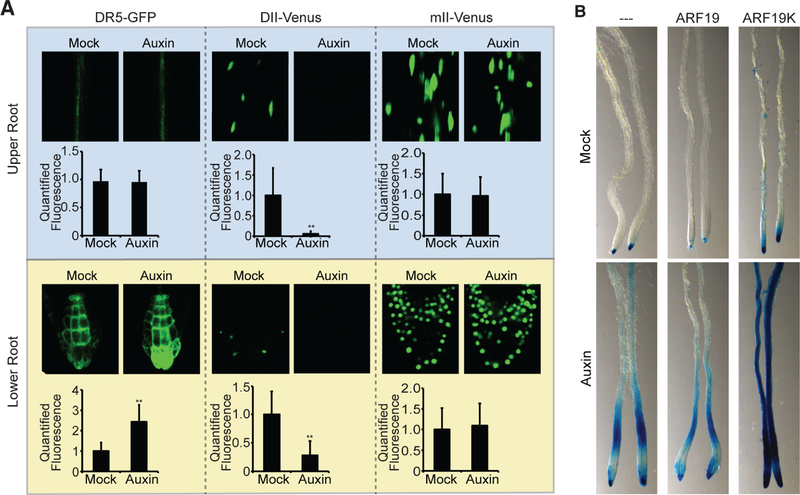
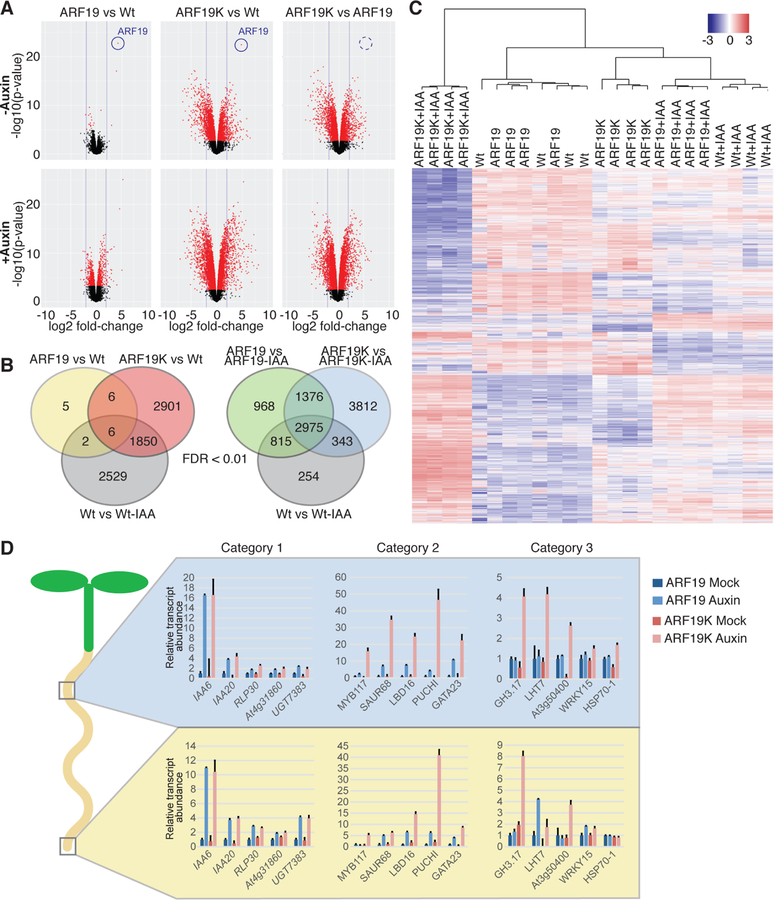
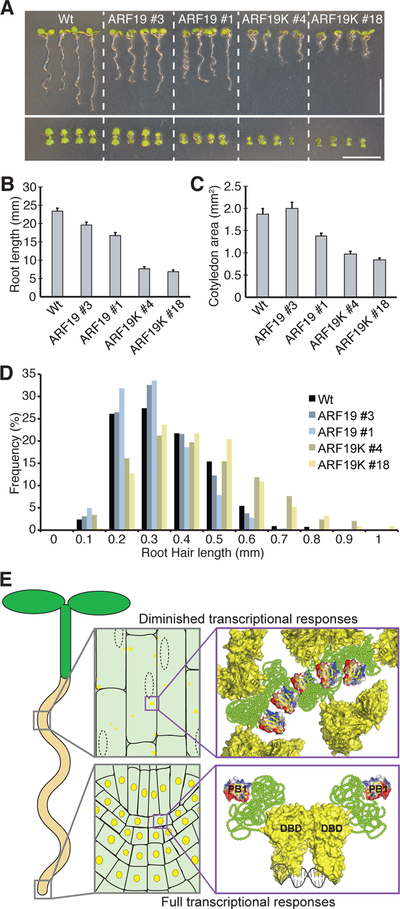
The phytohormone auxin plays crucial roles in nearly every aspect of plant growth and development. The auxin response factor (ARF) transcription factor family regulates auxin-responsive gene expression and exhibits nuclear localization in regions of high auxin responsiveness. Here we show that the ARF7 and ARF19 proteins accumulate in micron-sized assemblies within the cytoplasm of tissues with attenuated auxin responsiveness. We found that the intrinsically disordered middle region and the folded PB1 interaction domain of ARFs drive protein assembly formation. Mutation of a single lysine within the PB1 domain abrogates cytoplasmic assemblies, promotes ARF nuclear localization, and results in an altered transcriptome and morphological defects. Our data suggest a model in which ARF nucleo-cytoplasmic partitioning regulates auxin responsiveness, providing a mechanism for cellular competence for auxin signaling.
Copyright © 2019 Elsevier Inc. All rights reserved.
Conflict of interest statement
DECLARATION OF INTERESTS
The authors declare no competing interests.
Figures
Similar articles
-
MCTP controls nucleocytoplasmic partitioning of AUXIN RESPONSE FACTORs during lateral root development.Dev Cell. 2024 Dec 16;59(24):3229-3244.e5. doi: 10.1016/j.devcel.2024.09.026. Epub 2024 Oct 17. Dev Cell. 2024. PMID: 39423818
-
A role for auxin response factor 19 in auxin and ethylene signaling in Arabidopsis.Plant Physiol. 2006 Mar;140(3):899-908. doi: 10.1104/pp.105.070987. Epub 2006 Feb 3. Plant Physiol. 2006. PMID: 16461383 Free PMC article.
-
Functional genomic analysis of the AUXIN RESPONSE FACTOR gene family members in Arabidopsis thaliana: unique and overlapping functions of ARF7 and ARF19.Plant Cell. 2005 Feb;17(2):444-63. doi: 10.1105/tpc.104.028316. Epub 2005 Jan 19. Plant Cell. 2005. PMID: 15659631 Free PMC article.
-
Genetics of Aux/IAA and ARF action in plant growth and development.Plant Mol Biol. 2002 Jun-Jul;49(3-4):387-400. Plant Mol Biol. 2002. PMID: 12036262 Review.
-
Auxin response factors are keys to the many auxin doors.New Phytol. 2022 Jul;235(2):402-419. doi: 10.1111/nph.18159. Epub 2022 May 10. New Phytol. 2022. PMID: 35434800 Review.
Cited by
-
AUXIN RESPONSE FACTOR protein accumulation and function.Bioessays. 2023 Nov;45(11):e2300018. doi: 10.1002/bies.202300018. Epub 2023 Aug 16. Bioessays. 2023. PMID: 37584215 Free PMC article. Review.
-
Liquid-liquid phase separation in plants: Advances and perspectives from model species to crops.Plant Commun. 2024 Jan 8;5(1):100663. doi: 10.1016/j.xplc.2023.100663. Epub 2023 Jul 26. Plant Commun. 2024. PMID: 37496271 Free PMC article. Review.
-
Nucleocytoplasmic partitioning as a mechanism to regulate Arabidopsis signaling events.Curr Opin Cell Biol. 2021 Apr;69:136-141. doi: 10.1016/j.ceb.2021.01.006. Epub 2021 Feb 20. Curr Opin Cell Biol. 2021. PMID: 33618244 Free PMC article. Review.
-
An Introduction to the Stickers-and-Spacers Framework as Applied to Biomolecular Condensates.Methods Mol Biol. 2023;2563:95-116. doi: 10.1007/978-1-0716-2663-4_4. Methods Mol Biol. 2023. PMID: 36227469
-
Come together now: Dynamic body-formation of key regulators integrates environmental cues in plant development.Front Plant Sci. 2022 Nov 14;13:1052107. doi: 10.3389/fpls.2022.1052107. eCollection 2022. Front Plant Sci. 2022. PMID: 36452084 Free PMC article.
References
-
- Boer DR, Freire-Rios A, van den Berg WA, Saaki T, Manfield IW, Kepinski S, López-Vidrieo I, Franco-Zorrilla JM, de Vries SC, Solano R, et al. (2014). Structural Basis for DNA Binding Specificity by the Auxin-Dependent ARF Transcription Factors. Cell 156, 577–589. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
