

Natural selection contributed to immunological differences between hunter-gatherers and agriculturalists
- PMID: 31358949
- PMCID: PMC6684323
- DOI: 10.1038/s41559-019-0947-6
Natural selection contributed to immunological differences between hunter-gatherers and agriculturalists
Abstract
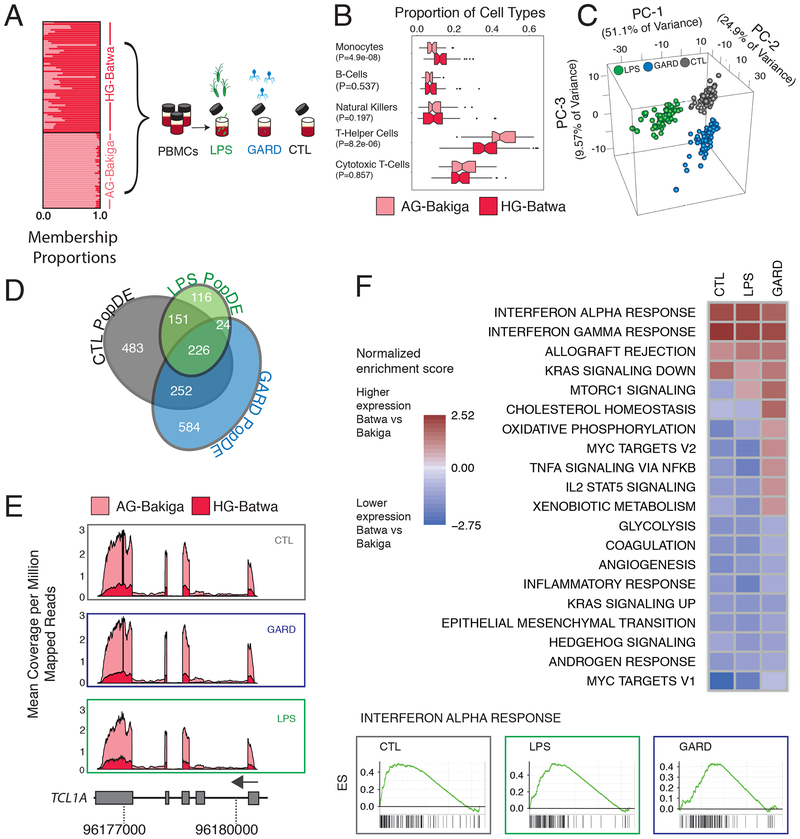
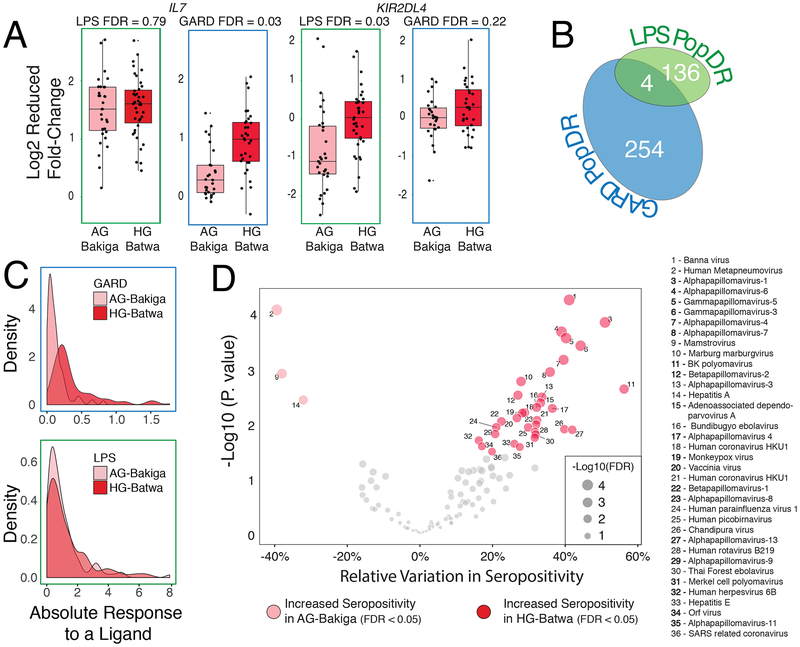
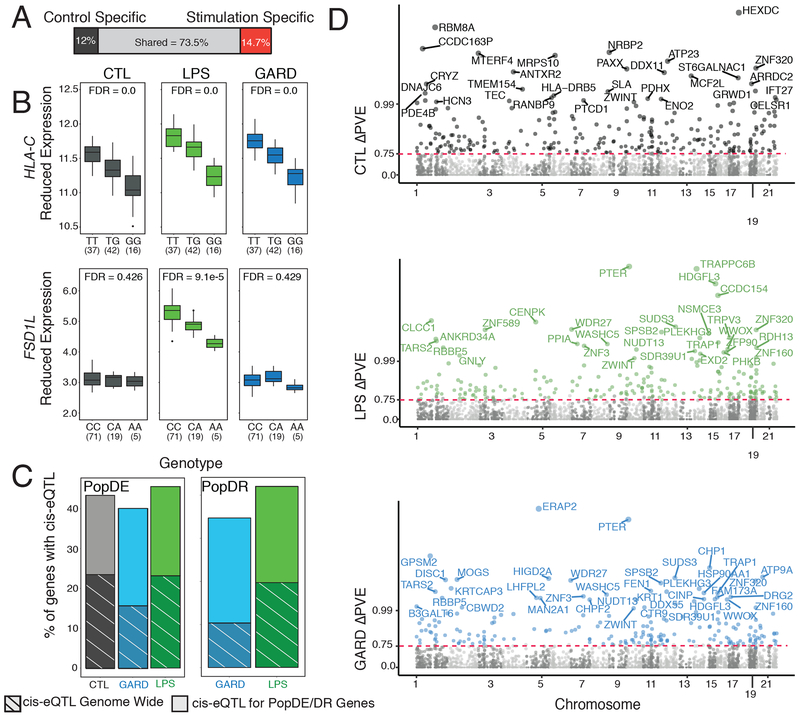
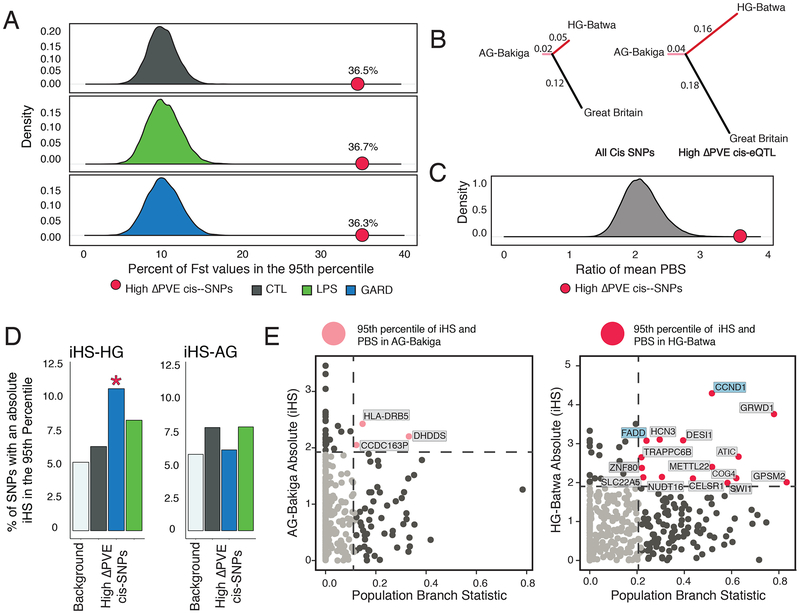
The shift from a hunter-gatherer to an agricultural mode of subsistence is believed to have been associated with profound changes in the burden and diversity of pathogens across human populations. Yet, the extent to which the advent of agriculture affected the evolution of the human immune system remains unknown. Here we present a comparative study of variation in the transcriptional responses of peripheral blood mononuclear cells to bacterial and viral stimuli between Batwa rainforest hunter-gatherers and Bakiga agriculturalists from Uganda. We observed increased divergence between hunter-gatherers and agriculturalists in the early transcriptional response to viruses compared with that for bacterial stimuli. We demonstrate that a significant fraction of these transcriptional differences are under genetic control and we show that positive natural selection has helped to shape population differences in immune regulation. Across the set of genetic variants underlying inter-population immune-response differences, however, the signatures of positive selection were disproportionately observed in the rainforest hunter-gatherers. This result is counter to expectations on the basis of the popularized notion that shifts in pathogen exposure due to the advent of agriculture imposed radically heightened selective pressures in agriculturalist populations.
Conflict of interest statement
Figures
Similar articles
-
Polygenic adaptation and convergent evolution on growth and cardiac genetic pathways in African and Asian rainforest hunter-gatherers.Proc Natl Acad Sci U S A. 2018 Nov 27;115(48):E11256-E11263. doi: 10.1073/pnas.1812135115. Epub 2018 Nov 9. Proc Natl Acad Sci U S A. 2018. PMID: 30413626 Free PMC article.
-
Adaptive, convergent origins of the pygmy phenotype in African rainforest hunter-gatherers.Proc Natl Acad Sci U S A. 2014 Sep 2;111(35):E3596-603. doi: 10.1073/pnas.1402875111. Epub 2014 Aug 18. Proc Natl Acad Sci U S A. 2014. PMID: 25136101 Free PMC article.
-
Recent Adaptive Acquisition by African Rainforest Hunter-Gatherers of the Late Pleistocene Sickle-Cell Mutation Suggests Past Differences in Malaria Exposure.Am J Hum Genet. 2019 Mar 7;104(3):553-561. doi: 10.1016/j.ajhg.2019.02.007. Epub 2019 Feb 28. Am J Hum Genet. 2019. PMID: 30827499 Free PMC article.
-
The demographic and adaptive history of central African hunter-gatherers and farmers.Curr Opin Genet Dev. 2018 Dec;53:90-97. doi: 10.1016/j.gde.2018.07.008. Epub 2018 Aug 10. Curr Opin Genet Dev. 2018. PMID: 30103089 Review.
-
Reconstructing prehistoric demography: What role for extant hunter-gatherers?Evol Anthropol. 2020 Nov;29(6):332-345. doi: 10.1002/evan.21869. Epub 2020 Oct 26. Evol Anthropol. 2020. PMID: 33103830 Review.
Cited by
-
Local genetic adaptation to habitat in wild chimpanzees.bioRxiv [Preprint]. 2024 Jul 9:2024.07.09.601734. doi: 10.1101/2024.07.09.601734. bioRxiv. 2024. PMID: 39026872 Free PMC article. Preprint.
-
DISSECT: deep semi-supervised consistency regularization for accurate cell type fraction and gene expression estimation.Genome Biol. 2024 Apr 30;25(1):112. doi: 10.1186/s13059-024-03251-5. Genome Biol. 2024. PMID: 38689377 Free PMC article.
-
Investigating the Influence of ANTXR2 Gene Mutations on Protective Antigen Binding for Heightened Anthrax Resistance.Genes (Basel). 2024 Mar 28;15(4):426. doi: 10.3390/genes15040426. Genes (Basel). 2024. PMID: 38674361 Free PMC article.
-
Epigenetic variation impacts individual differences in the transcriptional response to influenza infection.Nat Genet. 2024 Mar;56(3):408-419. doi: 10.1038/s41588-024-01668-z. Epub 2024 Feb 29. Nat Genet. 2024. PMID: 38424460
-
DNA methylation-environment interactions in the human genome.Elife. 2024 Feb 26;12:RP89371. doi: 10.7554/eLife.89371. Elife. 2024. PMID: 38407202 Free PMC article.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
