

The kinase PERK and the transcription factor ATF4 play distinct and essential roles in autophagy resulting from tunicamycin-induced ER stress
- PMID: 30926605
- PMCID: PMC6527152
- DOI: 10.1074/jbc.RA118.002829
The kinase PERK and the transcription factor ATF4 play distinct and essential roles in autophagy resulting from tunicamycin-induced ER stress
Abstract
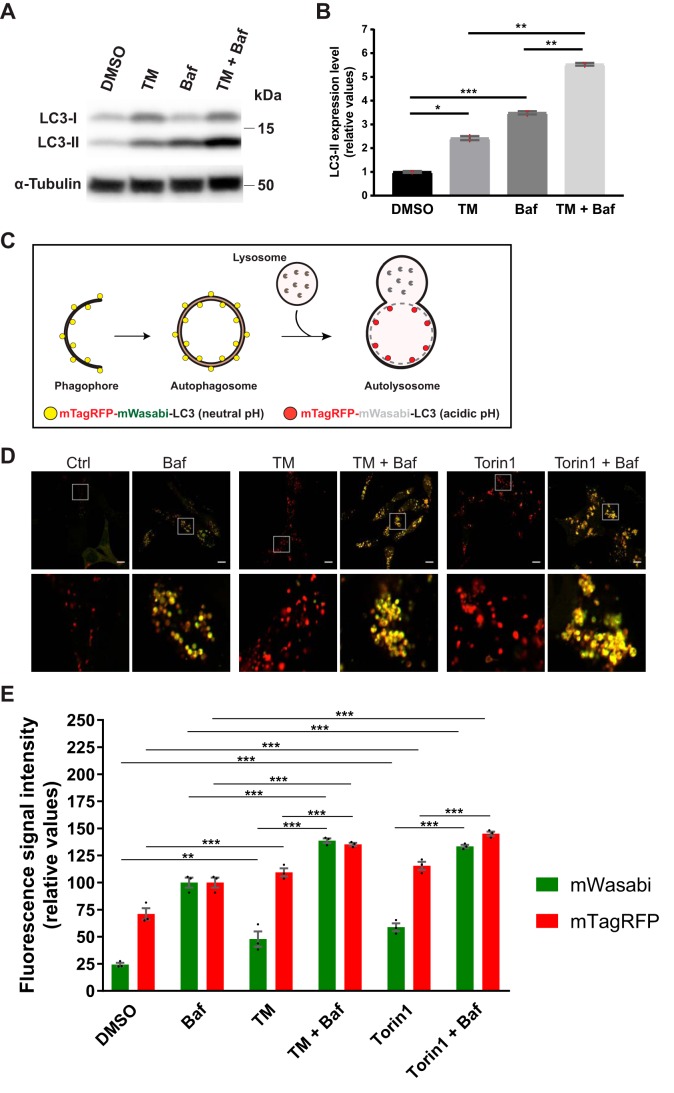
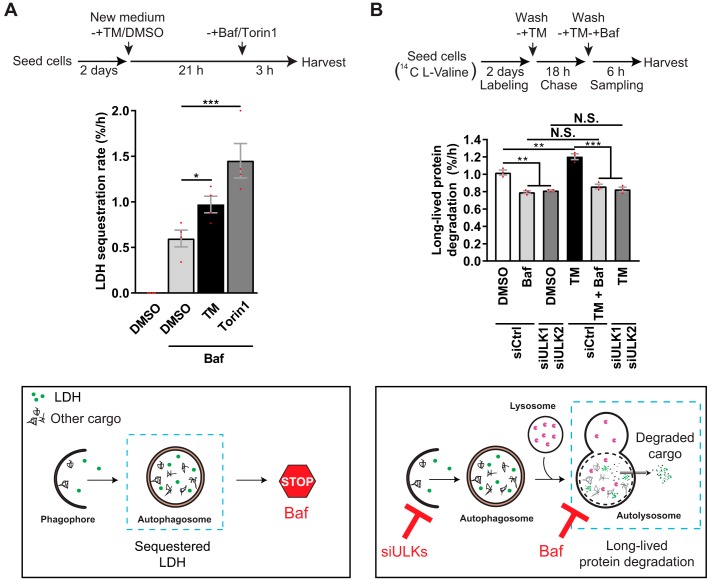
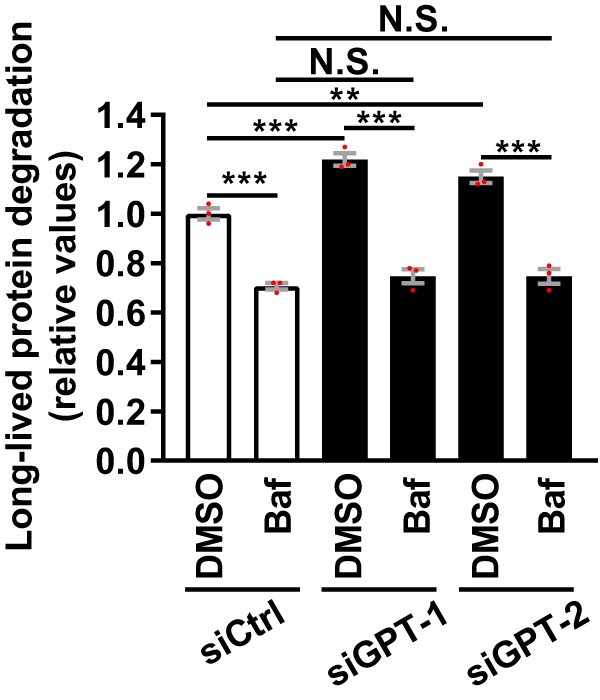
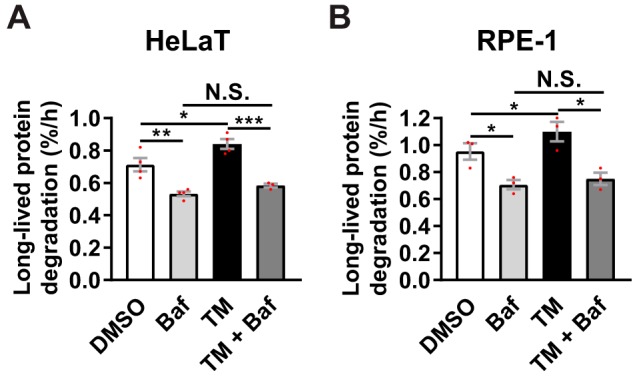
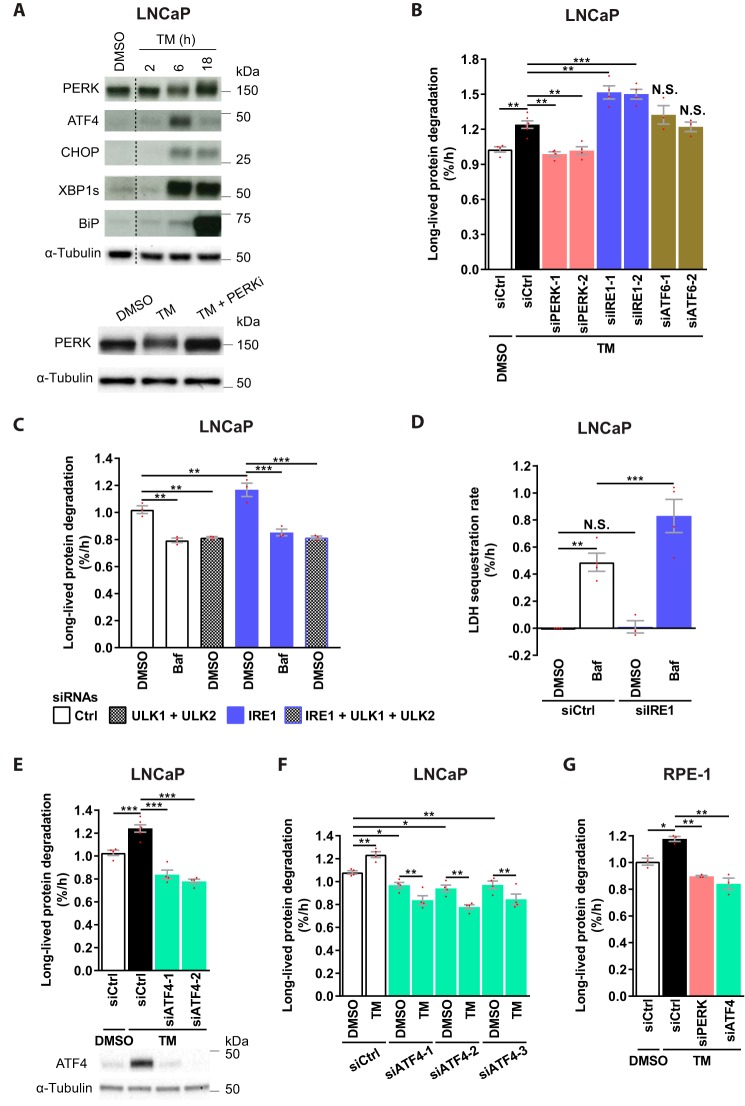
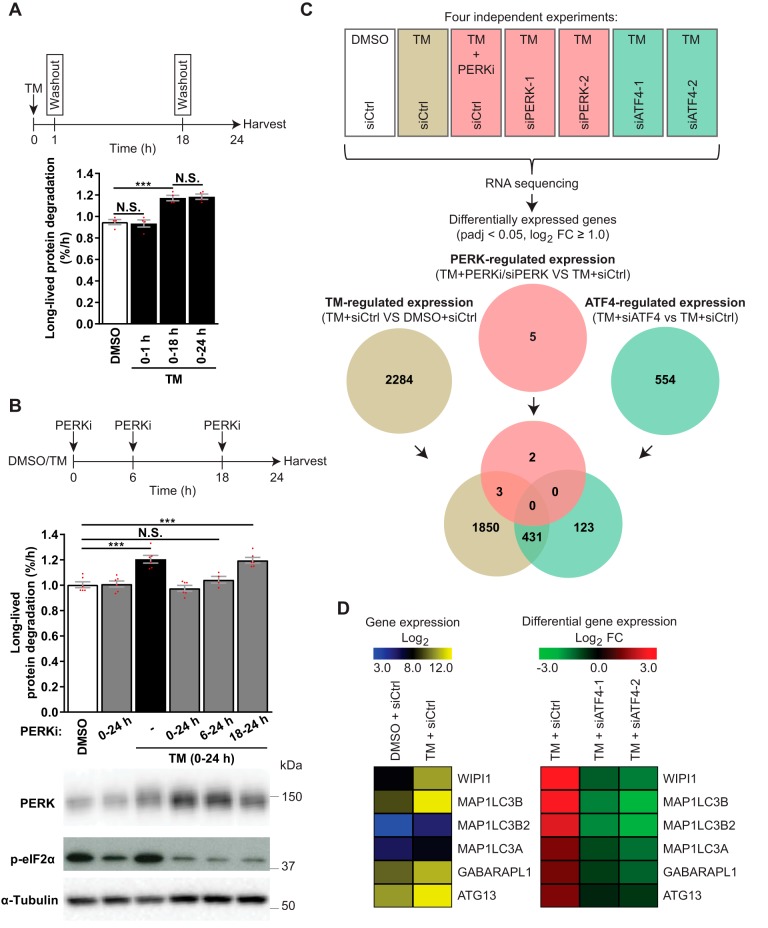
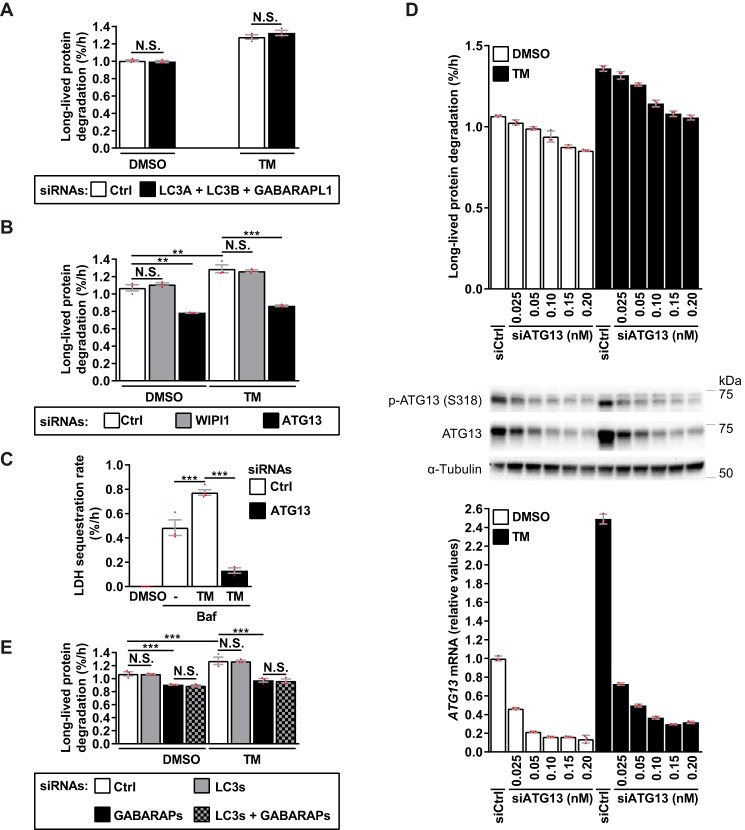
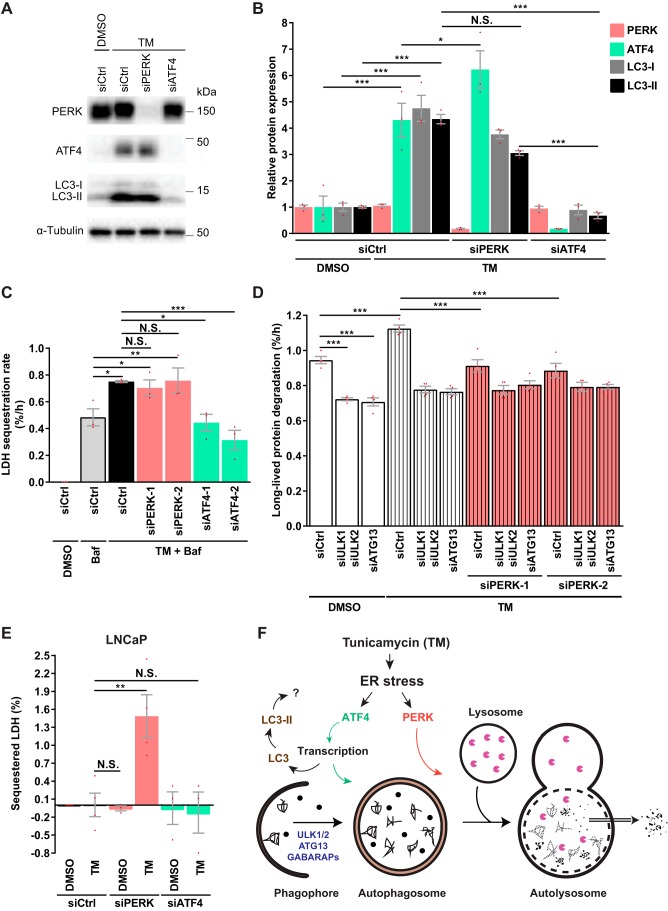
Endoplasmic reticulum (ER) stress is thought to activate autophagy via unfolded protein response (UPR)-mediated transcriptional up-regulation of autophagy machinery components and modulation of microtubule-associated protein 1 light chain 3 (LC3). The upstream UPR constituents pancreatic EIF2-α kinase (PERK) and inositol-requiring enzyme 1 (IRE1) have been reported to mediate these effects, suggesting that UPR may stimulate autophagy via PERK and IRE1. However, how the UPR and its components affect autophagic activity has not been thoroughly examined. By analyzing the flux of LC3 through the autophagic pathway, as well as the sequestration and degradation of autophagic cargo, we here conclusively show that the classical ER stressor tunicamycin (TM) enhances autophagic activity in mammalian cells. PERK and its downstream factor, activating transcription factor 4 (ATF4), were crucial for this induction, but surprisingly, IRE1 constitutively suppressed autophagic activity. TM-induced autophagy required autophagy-related 13 (ATG13), Unc-51-like autophagy-activating kinases 1/2 (ULK1/ULK2), and GABA type A receptor-associated proteins (GABARAPs), but interestingly, LC3 proteins appeared to be redundant. Strikingly, ATF4 was activated independently of PERK in both LNCaP and HeLa cells, and our further examination revealed that ATF4 and PERK regulated autophagy through separate mechanisms. Specifically, whereas ATF4 controlled transcription and was essential for autophagosome formation, PERK acted in a transcription-independent manner and was required at a post-sequestration step in the autophagic pathway. In conclusion, our results indicate that TM-induced UPR activates functional autophagy, and whereas IRE1 is a negative regulator, PERK and ATF4 are required at distinct steps in the autophagic pathway.
Keywords: GABA type A receptor-associated protein (GABARAP); activating transcription factor 4 (ATF4); autophagic degradation; autophagic sequestration; autophagy; endoplasmic reticulum stress (ER stress); inositol-requiring enzyme 1 (IRE1); microtubule-associated protein 1 light chain 3 (LC3); pancreatic EIF2-α kinase (PERK); protein degradation; signal transduction; tunicamycin (TM); unfolded protein response (UPR).
© 2019 Luhr et al.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
Similar articles
-
Cell death induced by the ER stressor thapsigargin involves death receptor 5, a non-autophagic function of MAP1LC3B, and distinct contributions from unfolded protein response components.Cell Commun Signal. 2020 Jan 27;18(1):12. doi: 10.1186/s12964-019-0499-z. Cell Commun Signal. 2020. PMID: 31987044 Free PMC article.
-
Dihydropyrazine induces endoplasmic reticulum stress and inhibits autophagy in HepG2 human hepatoma cells.J Toxicol Sci. 2024;49(7):313-319. doi: 10.2131/jts.49.313. J Toxicol Sci. 2024. PMID: 38945842
-
The Human Cytomegalovirus Endoplasmic Reticulum-Resident Glycoprotein UL148 Activates the Unfolded Protein Response.J Virol. 2018 Sep 26;92(20):e00896-18. doi: 10.1128/JVI.00896-18. Print 2018 Oct 15. J Virol. 2018. PMID: 30045994 Free PMC article.
-
The Role of the PERK/eIF2α/ATF4/CHOP Signaling Pathway in Tumor Progression During Endoplasmic Reticulum Stress.Curr Mol Med. 2016;16(6):533-44. doi: 10.2174/1566524016666160523143937. Curr Mol Med. 2016. PMID: 27211800 Free PMC article. Review.
-
Molecular Basis of Human Diseases and Targeted Therapy Based on Small-Molecule Inhibitors of ER Stress-Induced Signaling Pathways.Curr Mol Med. 2017;17(2):118-132. doi: 10.2174/1566524017666170306122643. Curr Mol Med. 2017. PMID: 28266275 Review.
Cited by
-
Detection of a Mitochondrial Fragmentation and Integrated Stress Response Using the Cell Painting Assay.J Med Chem. 2024 Aug 8;67(15):13252-13270. doi: 10.1021/acs.jmedchem.4c01183. Epub 2024 Jul 17. J Med Chem. 2024. PMID: 39018123 Free PMC article.
-
Endoplasmic reticulum stress-mediated cell death in cardiovascular disease.Cell Stress Chaperones. 2024 Feb;29(1):158-174. doi: 10.1016/j.cstres.2023.12.003. Epub 2024 Jan 29. Cell Stress Chaperones. 2024. PMID: 38295944 Free PMC article. Review.
-
The Role of Endoplasmic Reticulum Stress on Reducing Recombinant Protein Production in Mammalian Cells.Biochem Eng J. 2024 Oct;210:109434. doi: 10.1016/j.bej.2024.109434. Epub 2024 Jul 20. Biochem Eng J. 2024. PMID: 39220803
-
Effects of Spatial Expression of Activating Transcription Factor 4 on the Pathogenicity of Two Phenotypes of Bovine Viral Diarrhea Virus by Regulating the Endoplasmic Reticulum-Mediated Autophagy Process.Microbiol Spectr. 2023 Mar 20;11(2):e0422522. doi: 10.1128/spectrum.04225-22. Online ahead of print. Microbiol Spectr. 2023. PMID: 36939351 Free PMC article.
-
Regulation of autophagy by canonical and non-canonical ER stress responses.Semin Cancer Biol. 2020 Nov;66:116-128. doi: 10.1016/j.semcancer.2019.11.007. Epub 2019 Dec 12. Semin Cancer Biol. 2020. PMID: 31838023 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
