

Ubiquitin C-terminal hydrolase L1 (UCH-L1) loss causes neurodegeneration by altering protein turnover in the first postnatal weeks
- PMID: 30923110
- PMCID: PMC6475369
- DOI: 10.1073/pnas.1812413116
Ubiquitin C-terminal hydrolase L1 (UCH-L1) loss causes neurodegeneration by altering protein turnover in the first postnatal weeks
Abstract
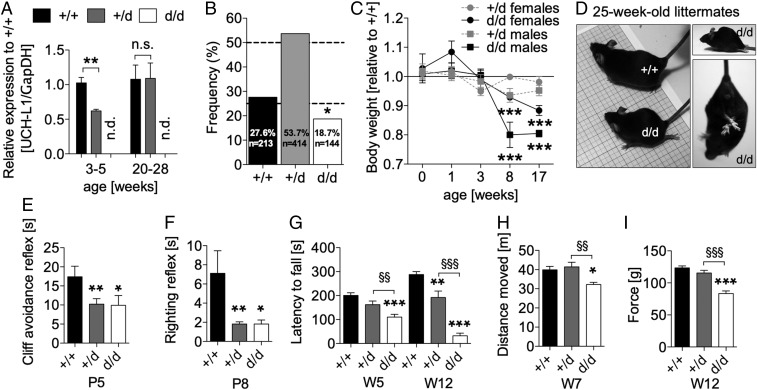
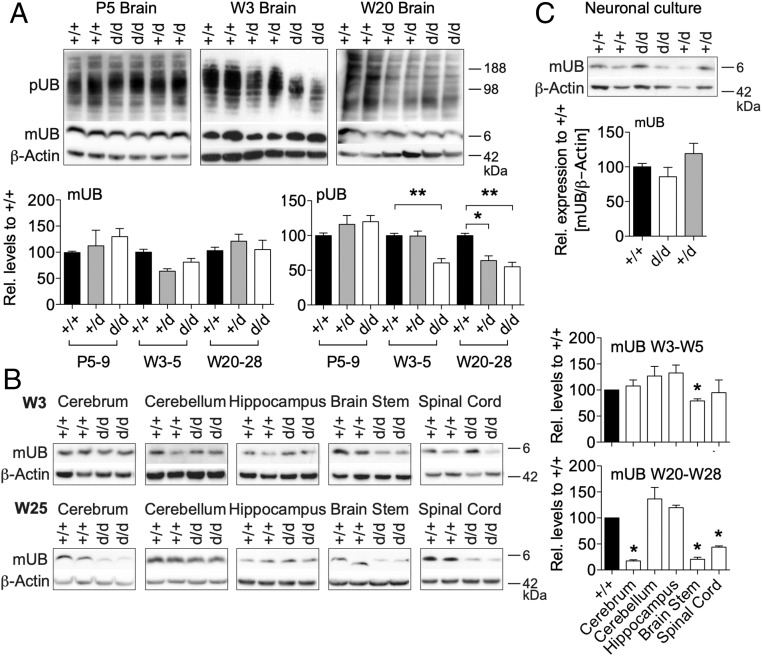
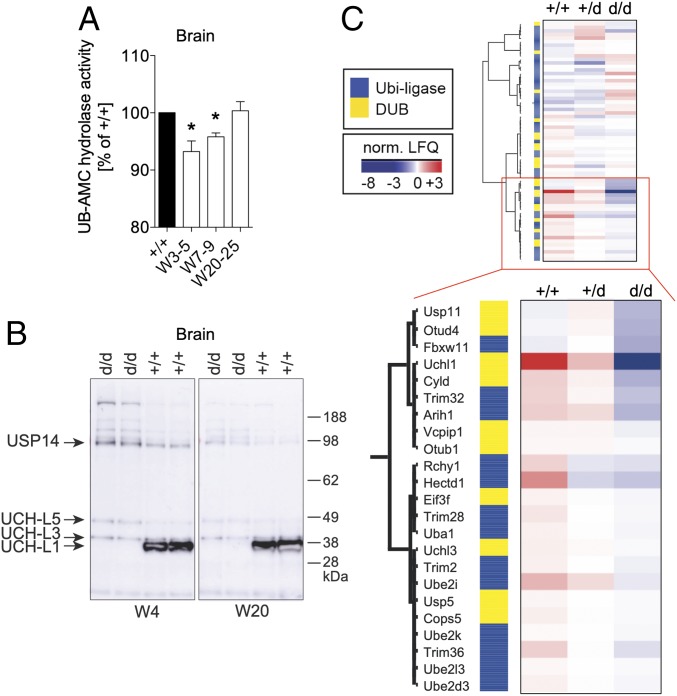
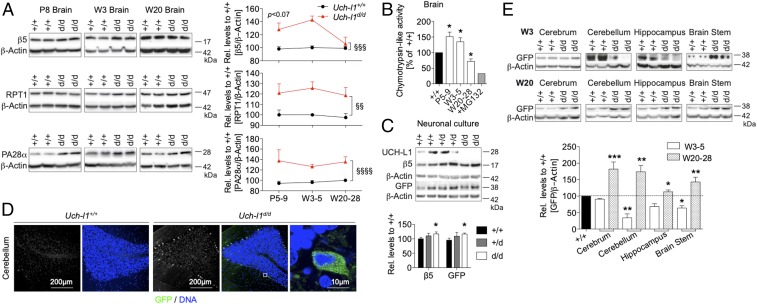
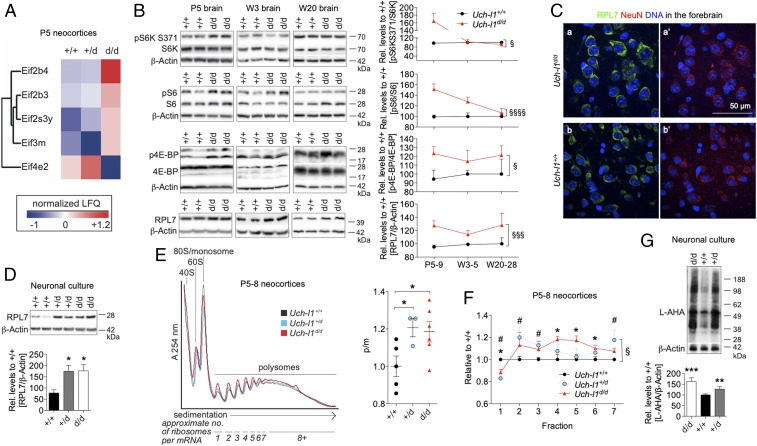
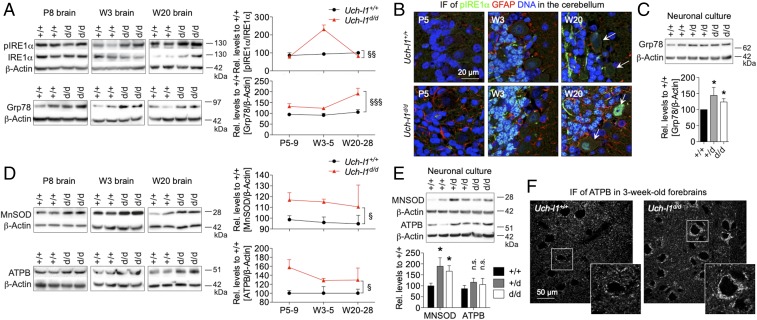
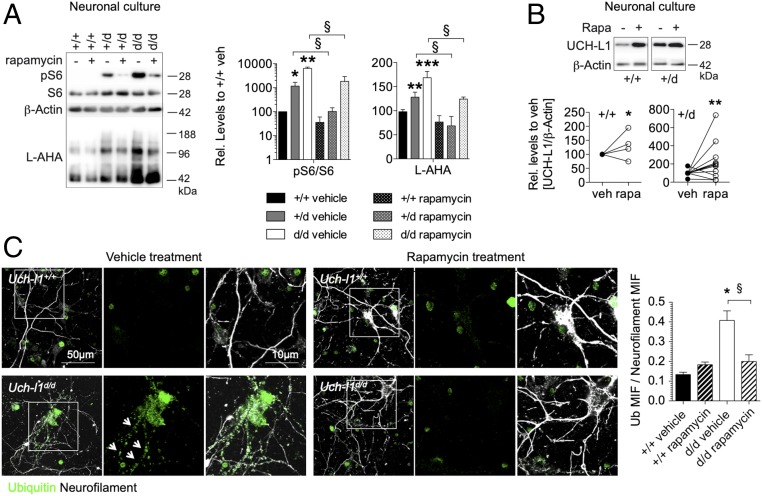
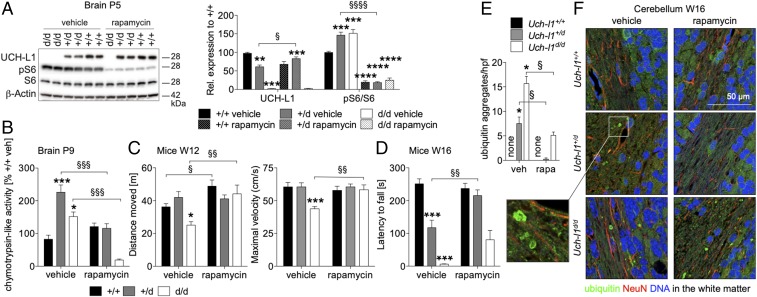
Ubiquitin C-terminal hydrolase L1 (UCH-L1) is one of the most abundant and enigmatic enzymes of the CNS. Based on existing UCH-L1 knockout models, UCH-L1 is thought to be required for the maintenance of axonal integrity, but not for neuronal development despite its high expression in neurons. Several lines of evidence suggest a role for UCH-L1 in mUB homeostasis, although the specific in vivo substrate remains elusive. Since the precise mechanisms underlying UCH-L1-deficient neurodegeneration remain unclear, we generated a transgenic mouse model of UCH-L1 deficiency. By performing biochemical and behavioral analyses we can show that UCH-L1 deficiency causes an acceleration of sensorimotor reflex development in the first postnatal week followed by a degeneration of motor function starting at periadolescence in the setting of normal cerebral mUB levels. In the first postnatal weeks, neuronal protein synthesis and proteasomal protein degradation are enhanced, with endoplasmic reticulum stress, and energy depletion, leading to proteasomal impairment and an accumulation of nondegraded ubiquitinated protein. Increased protein turnover is associated with enhanced mTORC1 activity restricted to the postnatal period in UCH-L1-deficient brains. Inhibition of mTORC1 with rapamycin decreases protein synthesis and ubiquitin accumulation in UCH-L1-deficient neurons. Strikingly, rapamycin treatment in the first 8 postnatal days ameliorates the neurological phenotype of UCH-L1-deficient mice up to 16 weeks, suggesting that early control of protein homeostasis is imperative for long-term neuronal survival. In summary, we identified a critical presymptomatic period during which UCH-L1-dependent enhanced protein synthesis results in neuronal strain and progressive loss of neuronal function.
Keywords: UCH-L1; development; mTOR; neurodegeneration; protein synthesis.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Ubiquitin C-Terminal Hydrolase L1 is required for regulated protein degradation through the ubiquitin proteasome system in kidney.Kidney Int. 2018 Jan;93(1):110-127. doi: 10.1016/j.kint.2017.05.016. Epub 2017 Jul 26. Kidney Int. 2018. PMID: 28754552
-
Ubiquitin C-terminal hydrolase-L1 protects cystic fibrosis transmembrane conductance regulator from early stages of proteasomal degradation.J Biol Chem. 2010 Apr 9;285(15):11314-25. doi: 10.1074/jbc.M109.044057. Epub 2010 Feb 10. J Biol Chem. 2010. PMID: 20147297 Free PMC article.
-
UCH-L1 induces podocyte hypertrophy in membranous nephropathy by protein accumulation.Biochim Biophys Acta. 2014 Jul;1842(7):945-58. doi: 10.1016/j.bbadis.2014.02.011. Epub 2014 Feb 28. Biochim Biophys Acta. 2014. PMID: 24583340
-
Ubiquitin C-terminal hydrolase L1 (UCH-L1): structure, distribution and roles in brain function and dysfunction.Biochem J. 2016 Aug 15;473(16):2453-62. doi: 10.1042/BCJ20160082. Biochem J. 2016. PMID: 27515257 Free PMC article. Review.
-
Ubiquitin C-terminal hydrolase-L1 (UCH-L1) as a therapeutic and diagnostic target in neurodegeneration, neurotrauma and neuro-injuries.Expert Opin Ther Targets. 2017 Jun;21(6):627-638. doi: 10.1080/14728222.2017.1321635. Epub 2017 Apr 27. Expert Opin Ther Targets. 2017. PMID: 28434268 Review.
Cited by
-
The spatial transcriptomic landscape of the healing mouse intestine following damage.Nat Commun. 2022 Feb 11;13(1):828. doi: 10.1038/s41467-022-28497-0. Nat Commun. 2022. PMID: 35149721 Free PMC article.
-
From Junk to Function: LncRNAs in CNS Health and Disease.Front Mol Neurosci. 2021 Jul 19;14:714768. doi: 10.3389/fnmol.2021.714768. eCollection 2021. Front Mol Neurosci. 2021. PMID: 34349622 Free PMC article. Review.
-
Abolishing UCHL1's hydrolase activity exacerbates TBI-induced axonal injury and neuronal death in mice.Exp Neurol. 2021 Feb;336:113524. doi: 10.1016/j.expneurol.2020.113524. Epub 2020 Nov 4. Exp Neurol. 2021. PMID: 33159930 Free PMC article.
-
The Ubiquitin Proteasome System in Neuromuscular Disorders: Moving Beyond Movement.Int J Mol Sci. 2020 Sep 3;21(17):6429. doi: 10.3390/ijms21176429. Int J Mol Sci. 2020. PMID: 32899400 Free PMC article. Review.
-
Abolishing UCHL1's hydrolase activity exacerbates ischemia-induced axonal injury and functional deficits in mice.J Cereb Blood Flow Metab. 2024 Nov;44(11):1349-1361. doi: 10.1177/0271678X241258809. Epub 2024 Jun 4. J Cereb Blood Flow Metab. 2024. PMID: 38833565 Free PMC article.
References
-
- Komander D, Clague MJ, Urbé S. Breaking the chains: Structure and function of the deubiquitinases. Nat Rev Mol Cell Biol. 2009;10:550–563. - PubMed
-
- Day IN, Thompson RJ. UCHL1 (PGP 9.5): Neuronal biomarker and ubiquitin system protein. Prog Neurobiol. 2010;90:327–362. - PubMed
-
- Wilkinson KD, et al. The neuron-specific protein PGP 9.5 is a ubiquitin carboxyl-terminal hydrolase. Science. 1989;246:670–673. - PubMed
-
- Meyer-Schwesinger C, et al. A new role for the neuronal ubiquitin C-terminal hydrolase-L1 (UCH-L1) in podocyte process formation and podocyte injury in human glomerulopathies. J Pathol. 2009;217:452–464. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
