

Inhibition of Influenza A Virus Replication by TRIM14 via Its Multifaceted Protein-Protein Interaction With NP
- PMID: 30873142
- PMCID: PMC6401474
- DOI: 10.3389/fmicb.2019.00344
Inhibition of Influenza A Virus Replication by TRIM14 via Its Multifaceted Protein-Protein Interaction With NP
Abstract
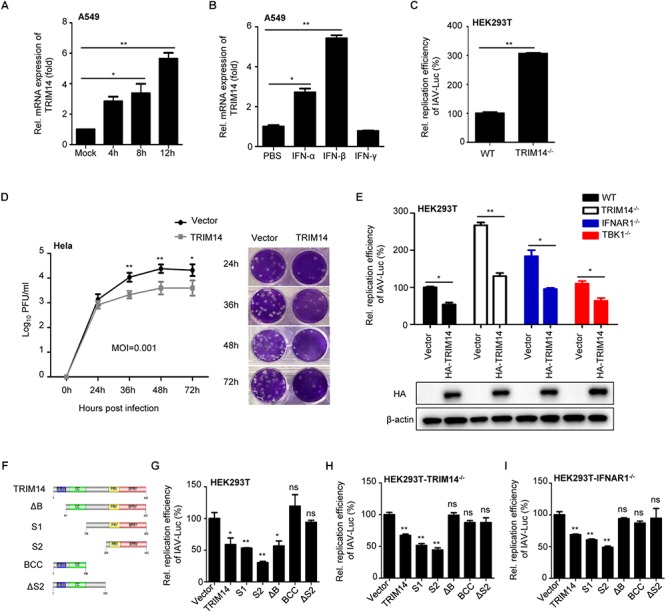
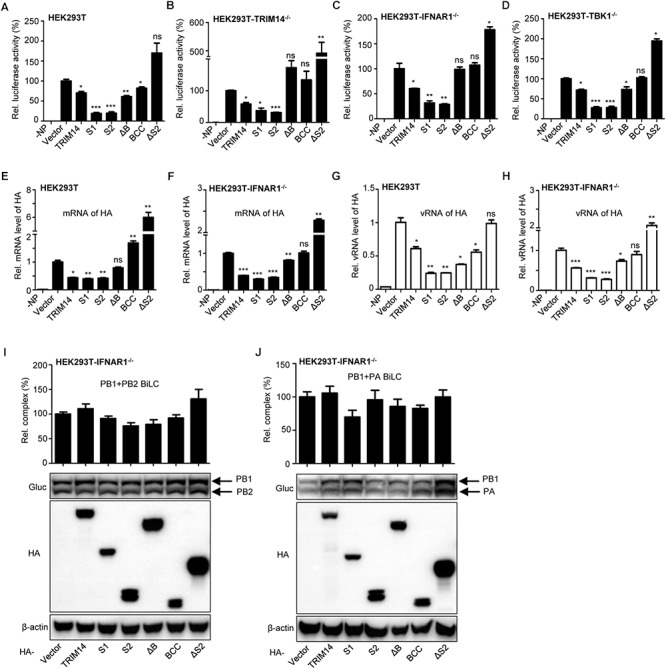
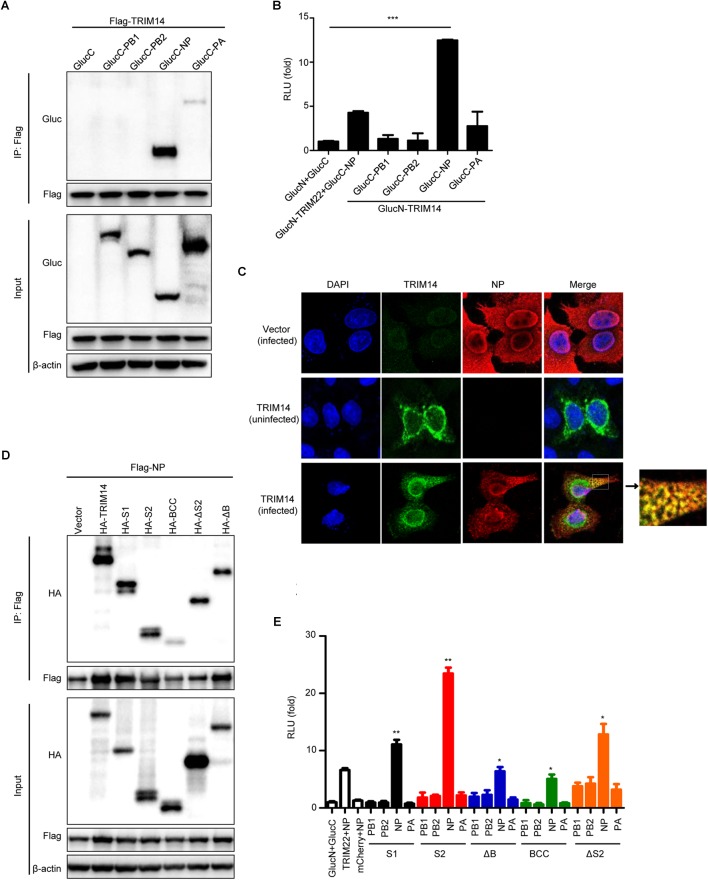
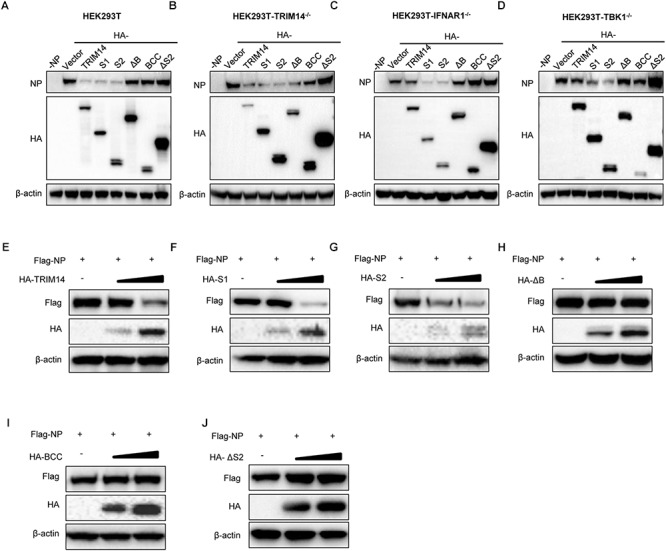
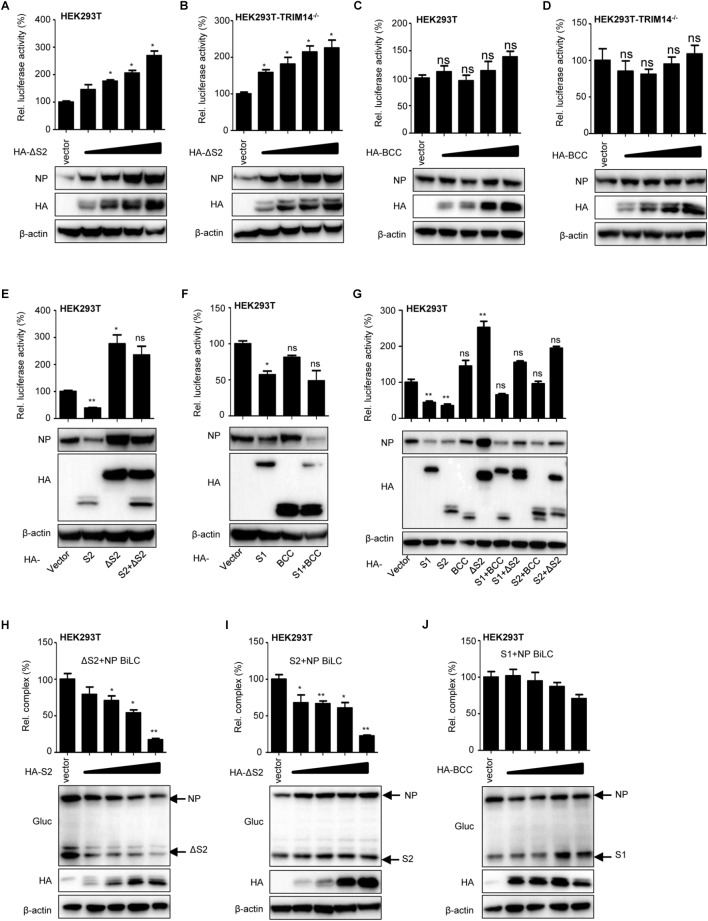
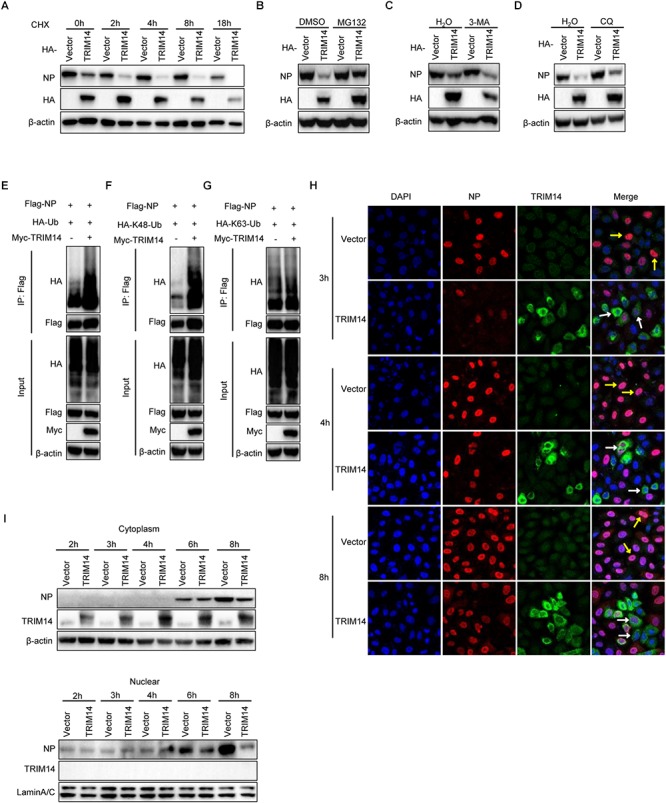
Influenza A virus (IAV) is a worldwide ongoing health threat causing diseases in both humans and animals. The interaction between IAV and host is a dynamic and evolving process that influences the pathogenicity and host specificity of the virus. TRIM14, a member of tripartite motif (TRIM) family, has been demonstrated to possess a strong capability of regulating type I interferon and NF-κB induction in host defense against viral infection. In this study, we found that TRIM14 could restrict the replication of IAV in a type I interferon and NF-κB independent manner. Mechanistically, different domains of TRIM14 could selectively interact with the viral nucleoprotein (NP), resulting in disparate influences on the RNP formation and viral replication. In particular, the PRYSPRY domain of TRIM14 exhibited a potent inhibitory activity on NP protein stability and IAV replication. On the contrary, the ΔS2 domain could rather antagonize the function of PRYSPRY domain and promote the IAV RNP formation by stabilizing NP. At the biochemical level, TRIM14-NP interaction could induce the K48-linked ubiquitination and proteasomal degradation of NP. Moreover, due to the rapid degradation of newly synthesized NP, TRIM14 could effectively block the translocation of NP from cytoplasm to nucleus thus further restrain the propagation of IAV in host cells. Taken together, our study has unraveled a previously unknown mechanism of TRIM14 mediated inhibition on RNP formation and influenza virus replication, and provides a new paradigm of complex and multifaceted host-pathogen interaction between ISG and viral protein.
Keywords: NP; PRYSPRY domain; TRIM14; influenza A virus; protein–protein interaction.
Figures
Similar articles
-
Human TRIM14 protects transgenic mice from influenza A viral infection without activation of other innate immunity pathways.Genes Immun. 2021 May;22(1):56-63. doi: 10.1038/s41435-021-00128-6. Epub 2021 Apr 16. Genes Immun. 2021. PMID: 33864033
-
TRIM41-Mediated Ubiquitination of Nucleoprotein Limits Influenza A Virus Infection.J Virol. 2018 Jul 31;92(16):e00905-18. doi: 10.1128/JVI.00905-18. Print 2018 Aug 15. J Virol. 2018. PMID: 29899090 Free PMC article.
-
The Nucleoprotein of H7N9 Influenza Virus Positively Regulates TRAF3-Mediated Innate Signaling and Attenuates Viral Virulence in Mice.J Virol. 2020 Nov 23;94(24):e01640-20. doi: 10.1128/JVI.01640-20. Print 2020 Nov 23. J Virol. 2020. PMID: 33028715 Free PMC article.
-
Autophagy Promotes Replication of Influenza A Virus In Vitro.J Virol. 2019 Feb 5;93(4):e01984-18. doi: 10.1128/JVI.01984-18. Print 2019 Feb 15. J Virol. 2019. PMID: 30541828 Free PMC article.
-
Modulation of Innate Immune Responses by the Influenza A NS1 and PA-X Proteins.Viruses. 2018 Dec 12;10(12):708. doi: 10.3390/v10120708. Viruses. 2018. PMID: 30545063 Free PMC article. Review.
Cited by
-
Tripartite motif-containing protein 46 accelerates influenza A H7N9 virus infection by promoting K48-linked ubiquitination of TBK1.Virol J. 2022 Nov 3;19(1):176. doi: 10.1186/s12985-022-01907-x. Virol J. 2022. PMID: 36329446 Free PMC article.
-
Tripartite motif‑containing 14 may aggravate cardiac hypertrophy via the AKT signalling pathway in neonatal rat cardiomyocytes and transgenic mice.Mol Med Rep. 2023 Sep;28(3):173. doi: 10.3892/mmr.2023.13060. Epub 2023 Jul 28. Mol Med Rep. 2023. PMID: 37503784 Free PMC article.
-
Expression Profiling of the Tripartite Motif Family Genes in Chronic Hepatitis C Patients.ACS Omega. 2023 Jul 7;8(28):25370-25377. doi: 10.1021/acsomega.3c02800. eCollection 2023 Jul 18. ACS Omega. 2023. PMID: 37483213 Free PMC article.
-
Human TRIM14 protects transgenic mice from influenza A viral infection without activation of other innate immunity pathways.Genes Immun. 2021 May;22(1):56-63. doi: 10.1038/s41435-021-00128-6. Epub 2021 Apr 16. Genes Immun. 2021. PMID: 33864033
-
To TRIM or not to TRIM: the balance of host-virus interactions mediated by the ubiquitin system.J Gen Virol. 2019 Dec;100(12):1641-1662. doi: 10.1099/jgv.0.001341. J Gen Virol. 2019. PMID: 31661051 Free PMC article.
References
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous