

Aedes aegypti AgBR1 antibodies modulate early Zika virus infection of mice
- PMID: 30858571
- PMCID: PMC6533137
- DOI: 10.1038/s41564-019-0385-x
Aedes aegypti AgBR1 antibodies modulate early Zika virus infection of mice
Abstract
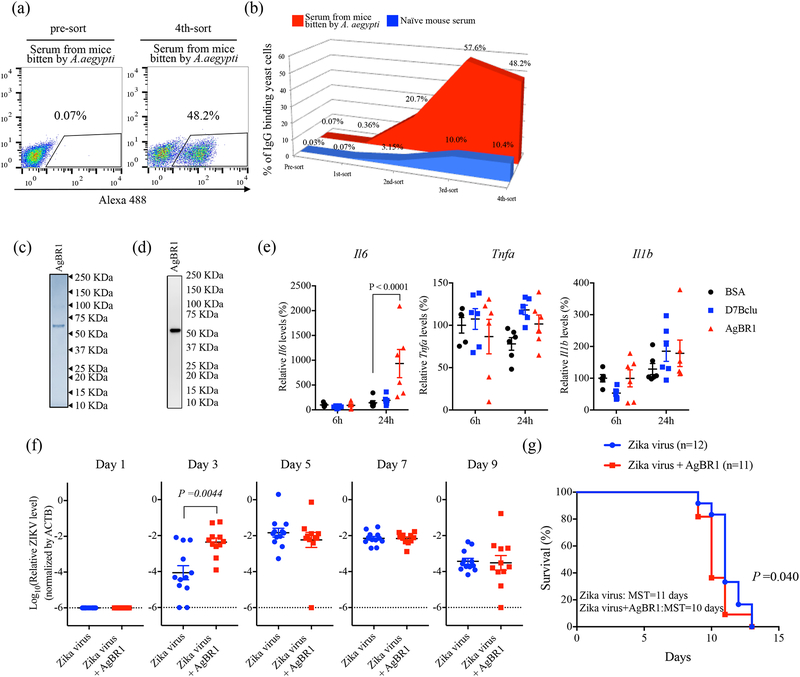
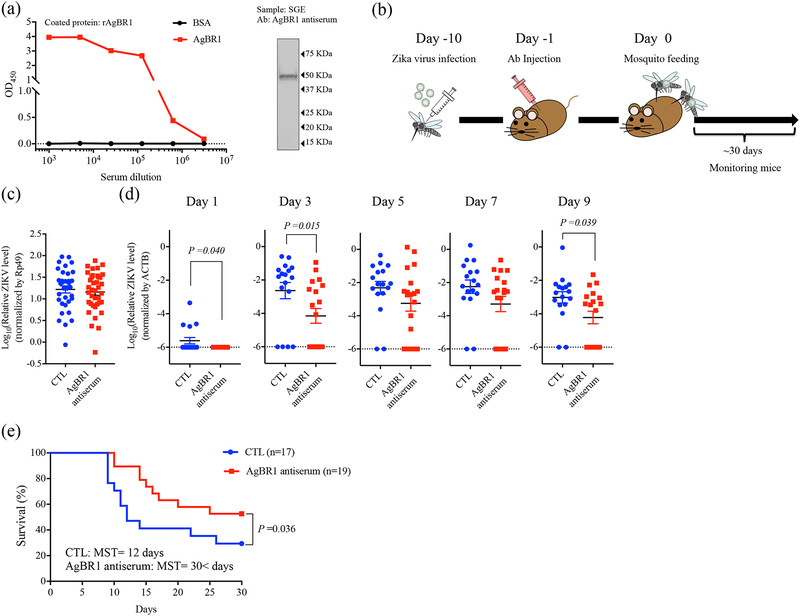
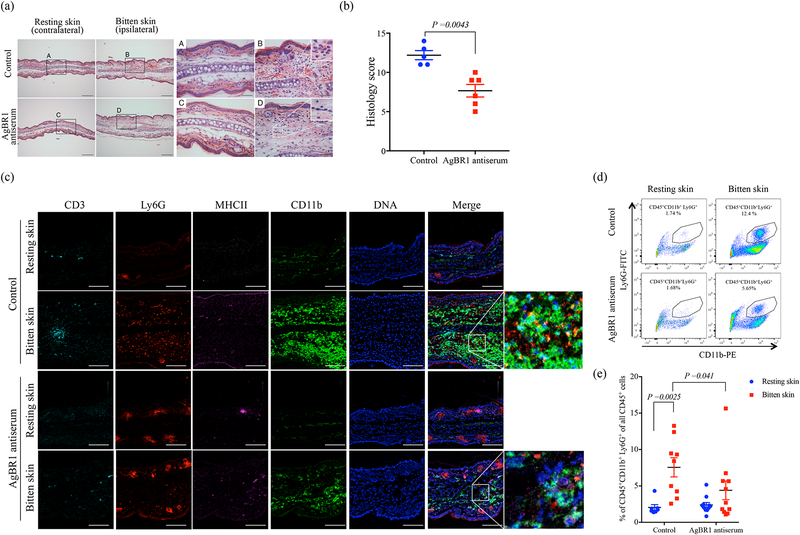
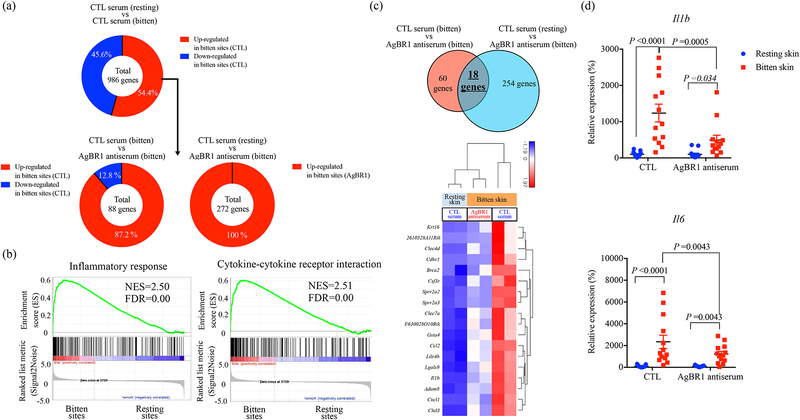
A recent epidemic of Zika virus in the Americas, affecting well over a million people, caused substantial mortality and morbidity, including Guillain-Barre syndrome, microcephaly and other fetal developmental defects1,2. Preventive and therapeutic measures that specifically target the virus are not readily available. The transmission of Zika virus is predominantly mosquito-borne, and Aedes aegypti mosquitoes serve as a key vector for Zika virus3. Here, to identify salivary factors that modulate mosquito-borne Zika virus infection, we focused on antigenic proteins in mice that were repeatedly bitten by mosquitoes and developed antibodies against salivary proteins. Using a yeast surface display screen, we identified five antigenic A. aegypti salivary proteins in mice. Antiserum against one of these five proteins-A. aegypti bacteria-responsive protein 1 (AgBR1)-suppressed early inflammatory responses in the skin of mice bitten by Zika-virus-infected mosquitoes. AgBR1 antiserum also partially protected mice from lethal mosquito-borne-but not needle-injected-Zika virus infection. These data suggest that AgBR1 is a target for the prevention of mosquito-transmitted Zika virus infection.
Conflict of interest statement
Competing Interests
The authors declare no competing interests.
Figures
Similar articles
-
Aedes aegypti NeSt1 Protein Enhances Zika Virus Pathogenesis by Activating Neutrophils.J Virol. 2019 Jun 14;93(13):e00395-19. doi: 10.1128/JVI.00395-19. Print 2019 Jul 1. J Virol. 2019. PMID: 30971475 Free PMC article.
-
AgBR1 and NeSt1 antisera protect mice from Aedes aegypti-borne Zika infection.Vaccine. 2021 Mar 19;39(12):1675-1679. doi: 10.1016/j.vaccine.2021.01.072. Epub 2021 Feb 20. Vaccine. 2021. PMID: 33622591 Free PMC article.
-
Zika virus transmission to mouse ear by mosquito bite: a laboratory model that replicates the natural transmission process.Parasit Vectors. 2017 Jul 20;10(1):346. doi: 10.1186/s13071-017-2286-2. Parasit Vectors. 2017. PMID: 28728607 Free PMC article.
-
The Convergence of a Virus, Mosquitoes, and Human Travel in Globalizing the Zika Epidemic.J Community Health. 2016 Jun;41(3):674-9. doi: 10.1007/s10900-016-0177-7. J Community Health. 2016. PMID: 26969497 Review.
-
Modes of Transmission of Zika Virus.J Infect Dis. 2017 Dec 16;216(suppl_10):S875-S883. doi: 10.1093/infdis/jix396. J Infect Dis. 2017. PMID: 29267909 Free PMC article. Review.
Cited by
-
Current Advances in Zika Vaccine Development.Vaccines (Basel). 2022 Oct 28;10(11):1816. doi: 10.3390/vaccines10111816. Vaccines (Basel). 2022. PMID: 36366325 Free PMC article. Review.
-
Zika Virus-A Reemerging Neurotropic Arbovirus Associated with Adverse Pregnancy Outcomes and Neuropathogenesis.Pathogens. 2024 Feb 15;13(2):177. doi: 10.3390/pathogens13020177. Pathogens. 2024. PMID: 38392915 Free PMC article. Review.
-
A mosquito salivary protein promotes flavivirus transmission by activation of autophagy.Nat Commun. 2020 Jan 14;11(1):260. doi: 10.1038/s41467-019-14115-z. Nat Commun. 2020. PMID: 31937766 Free PMC article.
-
The Effects of A Mosquito Salivary Protein on Sporozoite Traversal of Host Cells.J Infect Dis. 2021 Aug 2;224(3):544-553. doi: 10.1093/infdis/jiaa759. J Infect Dis. 2021. PMID: 33306099 Free PMC article.
-
Evaluation of cutaneous immune response in a controlled human in vivo model of mosquito bites.Nat Commun. 2022 Nov 17;13(1):7036. doi: 10.1038/s41467-022-34534-9. Nat Commun. 2022. PMID: 36396947 Free PMC article. Clinical Trial.
References
-
- Ventura CV, Maia M, Bravo-Filho V, Gois AL & Belfort R Jr. Zika virus in Brazil and macular atrophy in a child with microcephaly. Lancet 387, 228 (2016). - PubMed
-
- Olsen B & Lundkvist A [Zika virus - ancient virus gets new life in a new ecosystem. Microcephaly and Guillain-Barre syndrome are possible consequences when there is no background herd immunity in the population]. Lakartidningen 113(2016). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
