

Processing bodies control the selective translation for optimal development of Arabidopsis young seedlings
- PMID: 30850529
- PMCID: PMC6442596
- DOI: 10.1073/pnas.1900084116
Processing bodies control the selective translation for optimal development of Arabidopsis young seedlings
Erratum in
-
Correction for Jang et al., Processing bodies control the selective translation for optimal development of Arabidopsis young seedlings.Proc Natl Acad Sci U S A. 2019 Jun 18;116(25):12575. doi: 10.1073/pnas.1908132116. Epub 2019 Jun 10. Proc Natl Acad Sci U S A. 2019. PMID: 31182577 Free PMC article. No abstract available.
Abstract
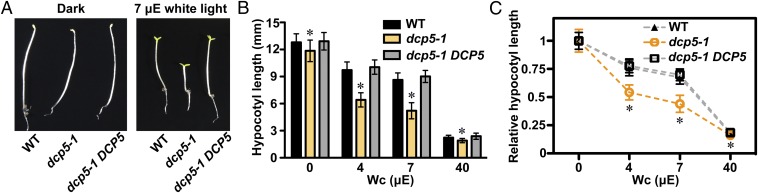
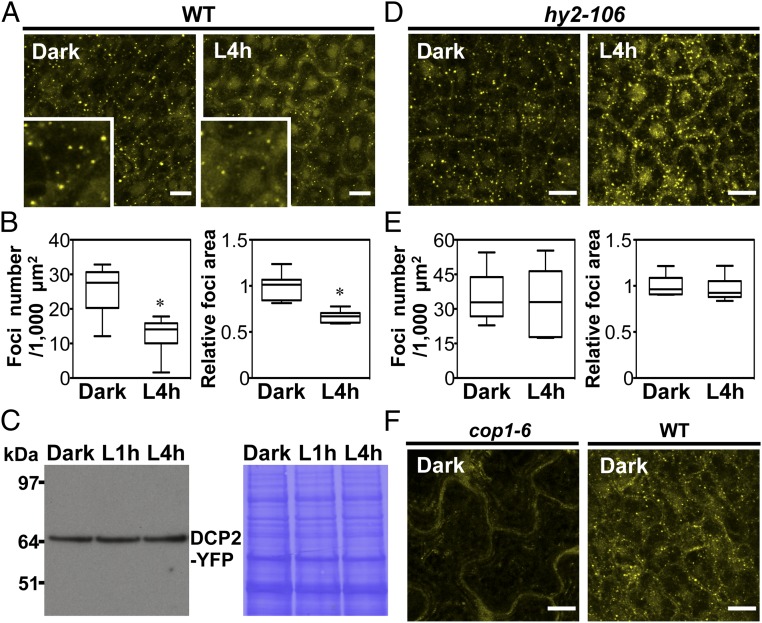
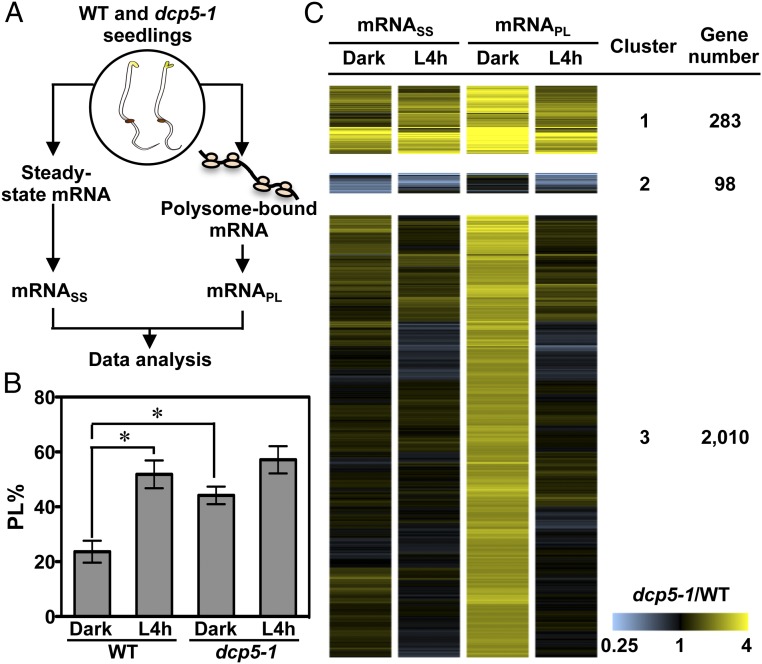
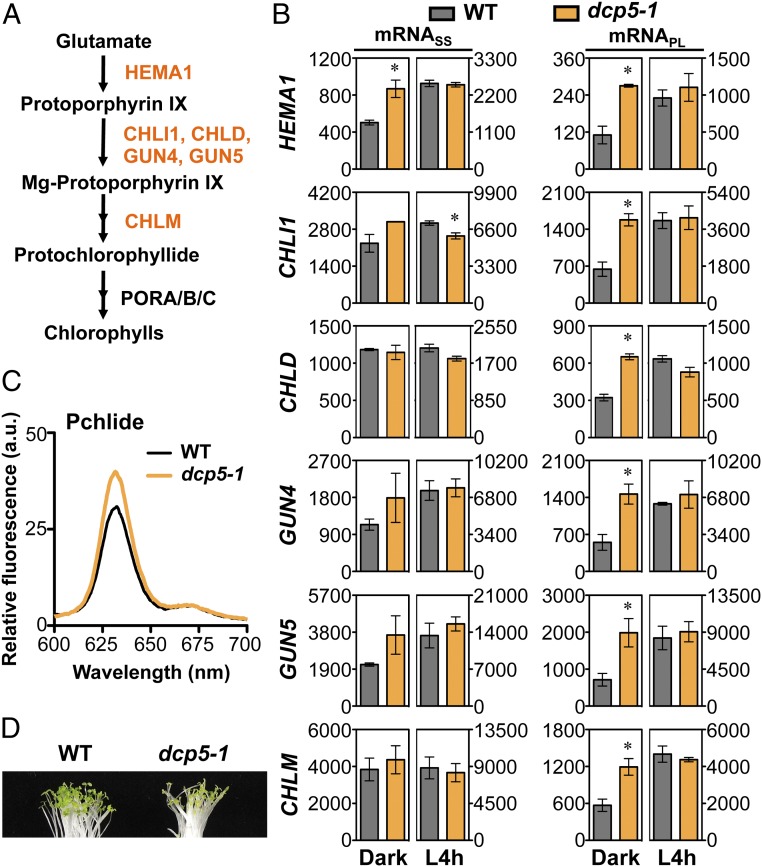
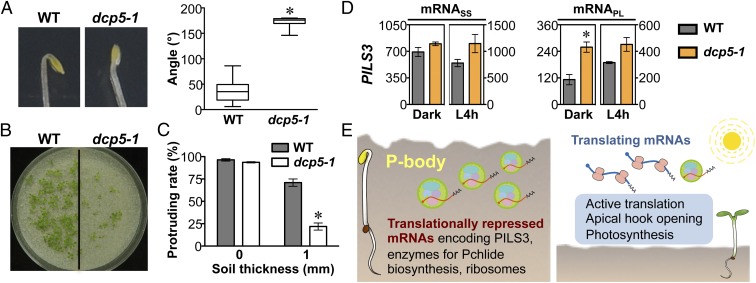
Germinated plant seeds buried in soil undergo skotomorphogenic development before emergence to reach the light environment. Young seedlings transitioning from dark to light undergo photomorphogenic development. During photomorphogenesis, light alters the transcriptome and enhances the translation of thousands of mRNAs during the dark-to-light transition in Arabidopsis young seedlings. About 1,500 of these mRNAs have comparable abundance before and after light treatment, which implies widespread translational repression in dark-grown seedlings. Processing bodies (p-bodies), the cytoplasmic granules found in diverse organisms, can balance the storage, degradation, and translation of mRNAs. However, the function of p-bodies in translation control remains largely unknown in plants. Here we found that an Arabidopsis mutant defective in p-body formation (Decapping 5; dcp5-1) showed reduced fitness under both dark and light conditions. Comparative transcriptome and translatome analyses of wild-type and dcp5-1 seedlings revealed that p-bodies can attenuate the premature translation of specific mRNAs in the dark, including those encoding enzymes for protochlorophyllide synthesis and PIN-LIKES3 for auxin-dependent apical hook opening. When the seedlings protrude from soil, light perception by photoreceptors triggers a reduced accumulation of p-bodies to release the translationally stalled mRNAs for active translation of mRNAs encoding proteins needed for photomorphogenesis. Our data support a key role for p-bodies in translation repression, an essential mechanism for proper skotomorphogenesis and timely photomorphogenesis in seedlings.
Keywords: light; p-bodies; photomorphogenesis; skotomorphogenesis; translation.
Copyright © 2019 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Definition of early transcriptional circuitry involved in light-induced reversal of PIF-imposed repression of photomorphogenesis in young Arabidopsis seedlings.Plant Cell. 2009 Nov;21(11):3535-53. doi: 10.1105/tpc.109.070672. Epub 2009 Nov 17. Plant Cell. 2009. PMID: 19920208 Free PMC article.
-
Arabidopsis decapping 5 is required for mRNA decapping, P-body formation, and translational repression during postembryonic development.Plant Cell. 2009 Oct;21(10):3270-9. doi: 10.1105/tpc.109.070078. Epub 2009 Oct 23. Plant Cell. 2009. PMID: 19855049 Free PMC article.
-
Arabidopsis DE-ETIOLATED1 represses photomorphogenesis by positively regulating phytochrome-interacting factors in the dark.Plant Cell. 2014 Sep;26(9):3630-45. doi: 10.1105/tpc.114.130666. Epub 2014 Sep 23. Plant Cell. 2014. PMID: 25248553 Free PMC article.
-
Gene expression regulation in photomorphogenesis from the perspective of the central dogma.Annu Rev Plant Biol. 2014;65:311-33. doi: 10.1146/annurev-arplant-050213-040337. Annu Rev Plant Biol. 2014. PMID: 24779996 Review.
-
To curve for survival: Apical hook development.J Integr Plant Biol. 2023 Feb;65(2):324-342. doi: 10.1111/jipb.13441. Epub 2023 Jan 31. J Integr Plant Biol. 2023. PMID: 36562414 Review.
Cited by
-
Get closer and make hotspots: liquid-liquid phase separation in plants.EMBO Rep. 2021 May 5;22(5):e51656. doi: 10.15252/embr.202051656. Epub 2021 Apr 28. EMBO Rep. 2021. PMID: 33913240 Free PMC article. Review.
-
APYRASE1/2 mediate red light-induced de-etiolation growth in Arabidopsis seedlings.Plant Physiol. 2022 Jun 27;189(3):1728-1740. doi: 10.1093/plphys/kiac150. Plant Physiol. 2022. PMID: 35357495 Free PMC article.
-
Plants use molecular mechanisms mediated by biomolecular condensates to integrate environmental cues with development.Plant Cell. 2023 Sep 1;35(9):3173-3186. doi: 10.1093/plcell/koad062. Plant Cell. 2023. PMID: 36879427 Free PMC article. Review.
-
A chemical genetic screen with the EXO70 inhibitor Endosidin2 uncovers potential modulators of exocytosis in Arabidopsis.Plant Direct. 2024 Jun 14;8(6):e592. doi: 10.1002/pld3.592. eCollection 2024 Jun. Plant Direct. 2024. PMID: 38881683 Free PMC article.
-
Dynamics of mRNA fate during light stress and recovery: from transcription to stability and translation.Plant J. 2024 Feb;117(3):818-839. doi: 10.1111/tpj.16531. Epub 2023 Nov 10. Plant J. 2024. PMID: 37947266 Free PMC article.
References
-
- Wu SH. Gene expression regulation in photomorphogenesis from the perspective of the central dogma. Annu Rev Plant Biol. 2014;65:311–333. - PubMed
-
- Xu X, Paik I, Zhu L, Huq E. Illuminating progress in phytochrome-mediated light signaling pathways. Trends Plant Sci. 2015;20:641–650. - PubMed
-
- Lau OS, Deng XW. The photomorphogenic repressors COP1 and DET1: 20 years later. Trends Plant Sci. 2012;17:584–593. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
