

Nucleotide Resolution Comparison of Transcription of Human Cytomegalovirus and Host Genomes Reveals Universal Use of RNA Polymerase II Elongation Control Driven by Dissimilar Core Promoter Elements
- PMID: 30755505
- PMCID: PMC6372792
- DOI: 10.1128/mBio.02047-18
Nucleotide Resolution Comparison of Transcription of Human Cytomegalovirus and Host Genomes Reveals Universal Use of RNA Polymerase II Elongation Control Driven by Dissimilar Core Promoter Elements
Abstract
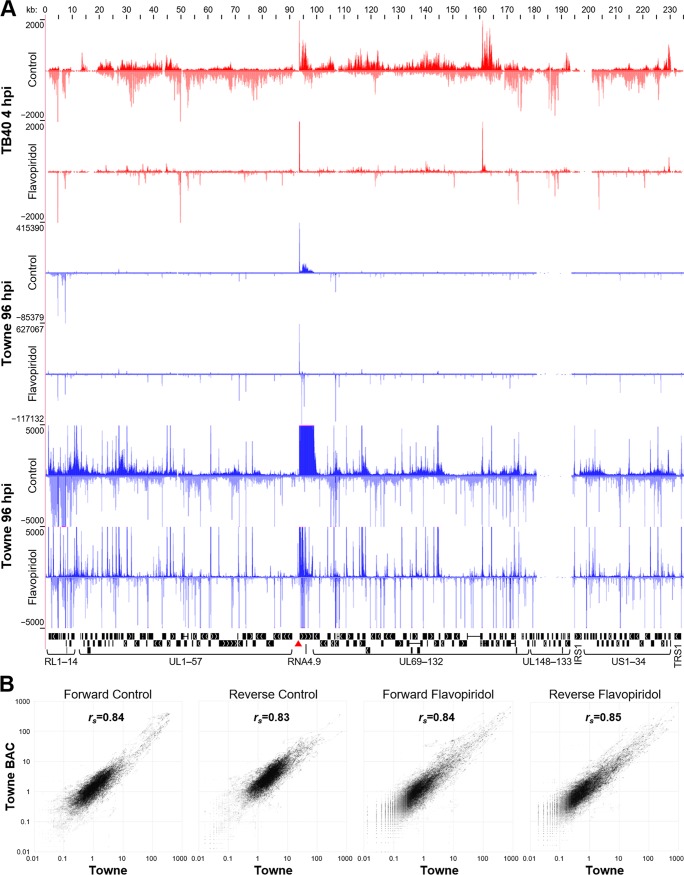
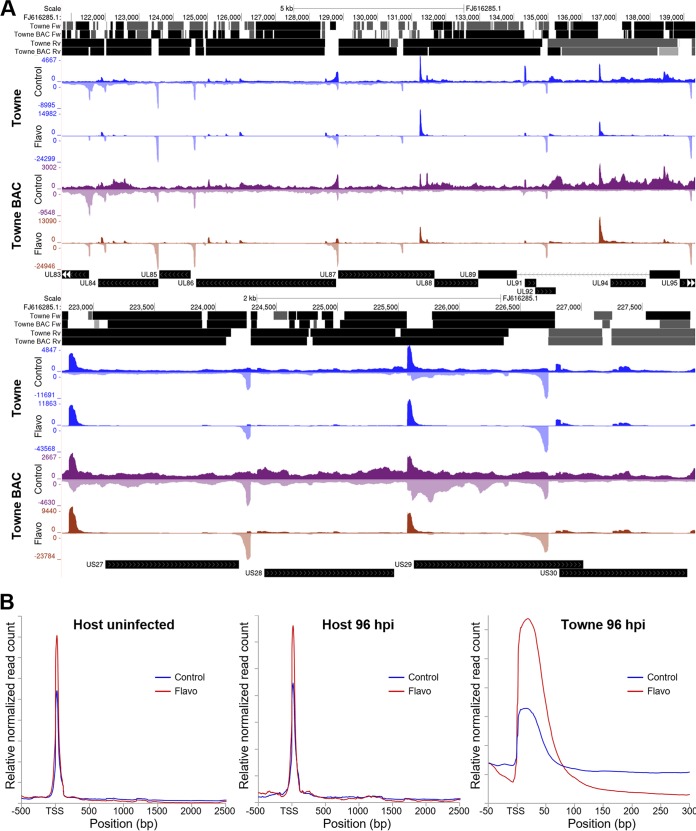
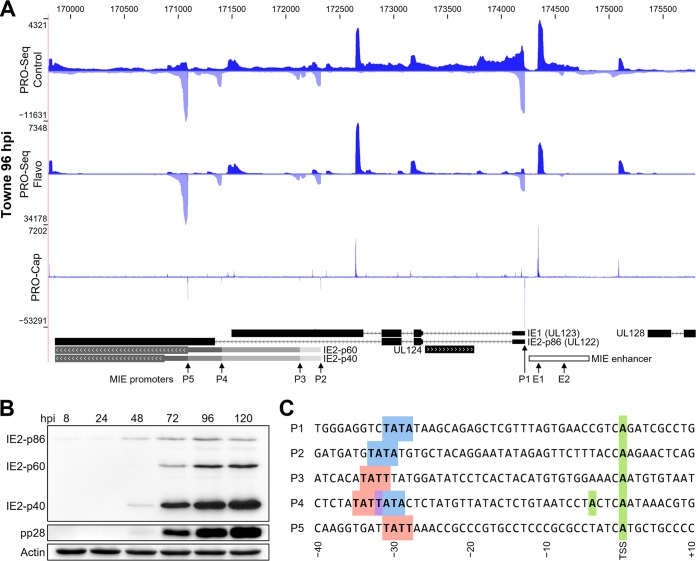
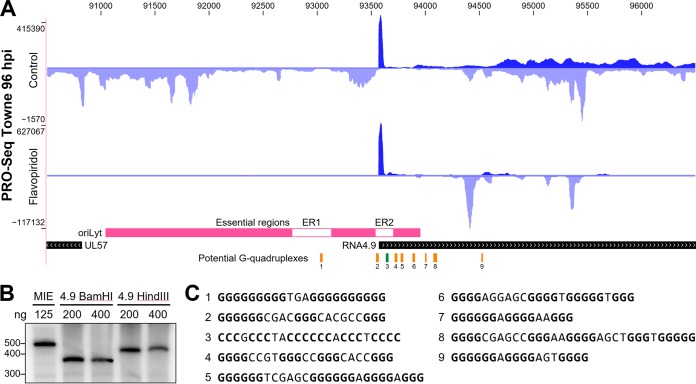
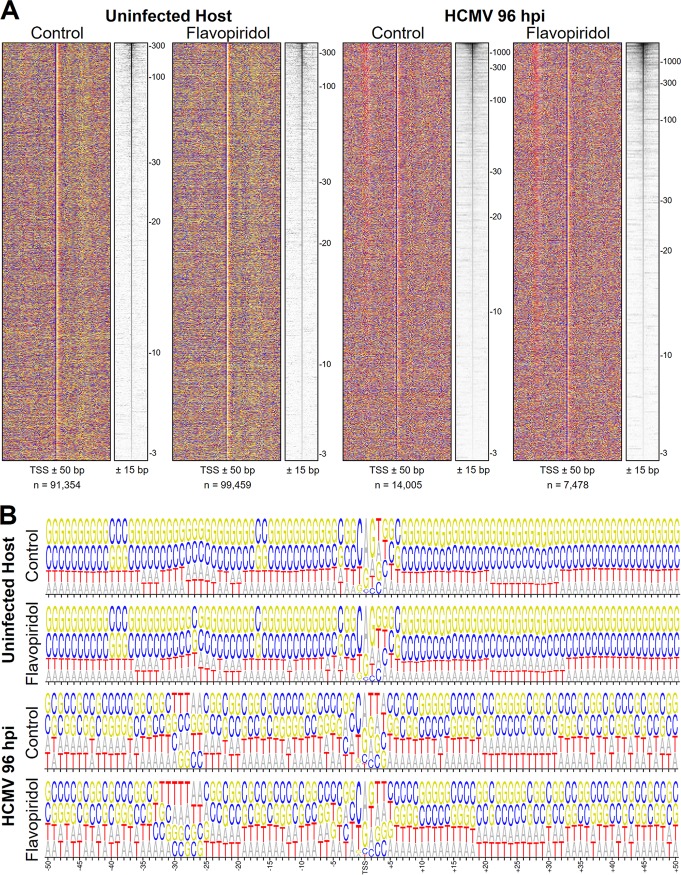
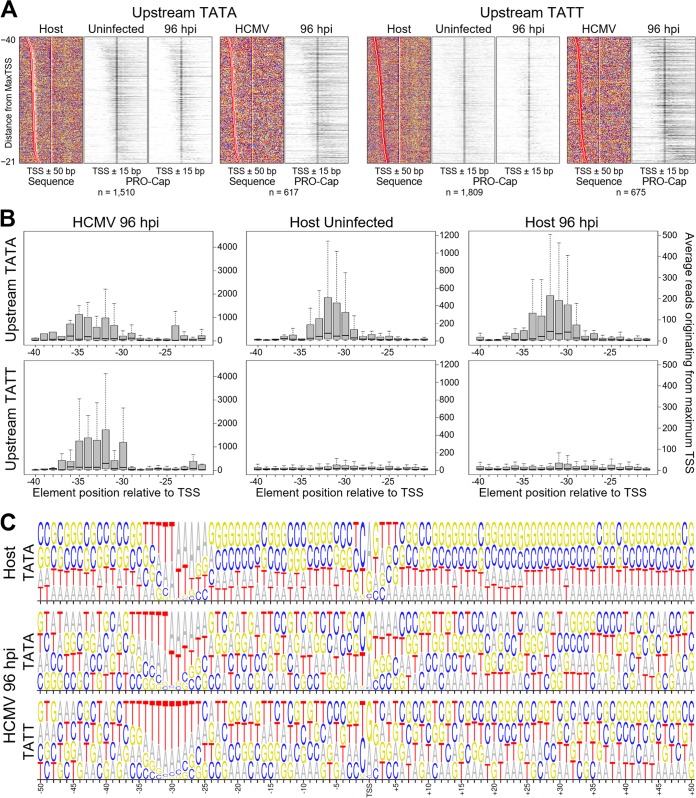
The large genome of human cytomegalovirus (HCMV) is transcribed by RNA polymerase II (Pol II). However, it is not known how closely this betaherpesvirus follows host transcriptional paradigms. We applied PRO-Seq and PRO-Cap methods to profile and quantify transcription initiation and productive elongation across the host and virus genomes in late infection. A major similarity between host transcription and viral transcription is that treatment of cells with the P-TEFb inhibitor flavopiridol preempts virtually all productive elongation, which otherwise covers most of the HCMV genome. The deep, nucleotide resolution identification of transcription start sites (TSSs) enabled an extensive analysis of core promoter elements. An important difference between host and viral transcription is that initiation is much more pervasive on the HCMV genome. The sequence preferences in the initiator region around the TSS and the utilization of upstream T/A-rich elements are different. Upstream TATA positions the TSS and boosts initiation in both the host and the virus, but upstream TATT has a significant stimulatory impact only on the viral template. The major immediate early (MIE) promoter remained active during late infection and was accompanied by transcription of both strands of the MIE enhancer from promoters within the enhancer. Surprisingly, we found that the long noncoding RNA4.9 is intimately associated with the viral origin of replication (oriLyt) and was transcribed to a higher level than any other viral or host promoter. Finally, our results significantly contribute to the idea that late in infection, transcription takes place on viral genomes that are not highly chromatinized.IMPORTANCE Human cytomegalovirus infects more than half of humans, persists silently in virtually all tissues, and produces life-threatening disease in immunocompromised individuals. HCMV is also the most common infectious cause of birth defects and the leading nongenetic cause of sensorineural hearing loss in the United States. Because there is no vaccine and current drugs have problems with potency, toxicity, and antiviral drug resistance, alternative treatment strategies that target different points of viral control are needed. Our current study contributes to this goal by applying newly developed methods to examine transcription of the HCMV and host genomes at nucleotide resolution in an attempt to find targetable differences between the two. After a thorough analysis of productive elongation and of core promoter element usage, we found that some mechanisms of regulating transcription are shared between the host and HCMV but that others are distinctly different. This suggests that HCMV transcription may be a legitimate target for future antiviral therapies and this might translate to other herpesviruses.
Keywords: P-TEFb; PRO-Seq; RNA polymerase II; RNA4.9; core promoter elements; cytomegalovirus.
Copyright © 2019 Parida et al.
Figures
Similar articles
-
Human cytomegalovirus IE2 drives transcription initiation from a select subset of late infection viral promoters by host RNA polymerase II.PLoS Pathog. 2020 Apr 6;16(4):e1008402. doi: 10.1371/journal.ppat.1008402. eCollection 2020 Apr. PLoS Pathog. 2020. PMID: 32251483 Free PMC article.
-
Cytomegalovirus late transcription factor target sequence diversity orchestrates viral early to late transcription.PLoS Pathog. 2021 Aug 2;17(8):e1009796. doi: 10.1371/journal.ppat.1009796. eCollection 2021 Aug. PLoS Pathog. 2021. PMID: 34339482 Free PMC article.
-
Human Cytomegalovirus IE2 Both Activates and Represses Initiation and Modulates Elongation in a Context-Dependent Manner.mBio. 2022 Jun 28;13(3):e0033722. doi: 10.1128/mbio.00337-22. Epub 2022 May 17. mBio. 2022. PMID: 35579393 Free PMC article.
-
Control of Immediate Early Gene Expression for Human Cytomegalovirus Reactivation.Front Cell Infect Microbiol. 2020 Sep 17;10:476. doi: 10.3389/fcimb.2020.00476. eCollection 2020. Front Cell Infect Microbiol. 2020. PMID: 33072616 Free PMC article. Review.
-
Core promoter-selective RNA polymerase II transcription.Biochem Soc Symp. 2006;(73):225-36. doi: 10.1042/bss0730225. Biochem Soc Symp. 2006. PMID: 16626302 Review.
Cited by
-
Reactivation of Epstein-Barr Virus from Latency Involves Increased RNA Polymerase Activity at CTCF Binding Sites on the Viral Genome.J Virol. 2023 Feb 28;97(2):e0189422. doi: 10.1128/jvi.01894-22. Epub 2023 Feb 6. J Virol. 2023. PMID: 36744959 Free PMC article.
-
The landscape of transcription initiation across latent and lytic KSHV genomes.PLoS Pathog. 2019 Jun 12;15(6):e1007852. doi: 10.1371/journal.ppat.1007852. eCollection 2019 Jun. PLoS Pathog. 2019. PMID: 31188901 Free PMC article.
-
Human cytomegalovirus IE2 drives transcription initiation from a select subset of late infection viral promoters by host RNA polymerase II.PLoS Pathog. 2020 Apr 6;16(4):e1008402. doi: 10.1371/journal.ppat.1008402. eCollection 2020 Apr. PLoS Pathog. 2020. PMID: 32251483 Free PMC article.
-
Cytomegalovirus late transcription factor target sequence diversity orchestrates viral early to late transcription.PLoS Pathog. 2021 Aug 2;17(8):e1009796. doi: 10.1371/journal.ppat.1009796. eCollection 2021 Aug. PLoS Pathog. 2021. PMID: 34339482 Free PMC article.
-
Cytomegalovirus Latency and Reactivation: An Intricate Interplay With the Host Immune Response.Front Cell Infect Microbiol. 2020 Mar 31;10:130. doi: 10.3389/fcimb.2020.00130. eCollection 2020. Front Cell Infect Microbiol. 2020. PMID: 32296651 Free PMC article. Review.
References
-
- Steurer B, Janssens RC, Geverts B, Geijer ME, Wienholz F, Theil AF, Chang J, Dealy S, Pothof J, van Cappellen WA, Houtsmuller AB, Marteijn JA. 2018. Live-cell analysis of endogenous GFP-RPB1 uncovers rapid turnover of initiating and promoter-paused RNA polymerase II. Proc Natl Acad Sci U S A 115:E4368–E4376. doi:10.1073/pnas.1717920115. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
