

TRIM2, a novel member of the antiviral family, limits New World arenavirus entry
- PMID: 30726215
- PMCID: PMC6380604
- DOI: 10.1371/journal.pbio.3000137
TRIM2, a novel member of the antiviral family, limits New World arenavirus entry
Abstract
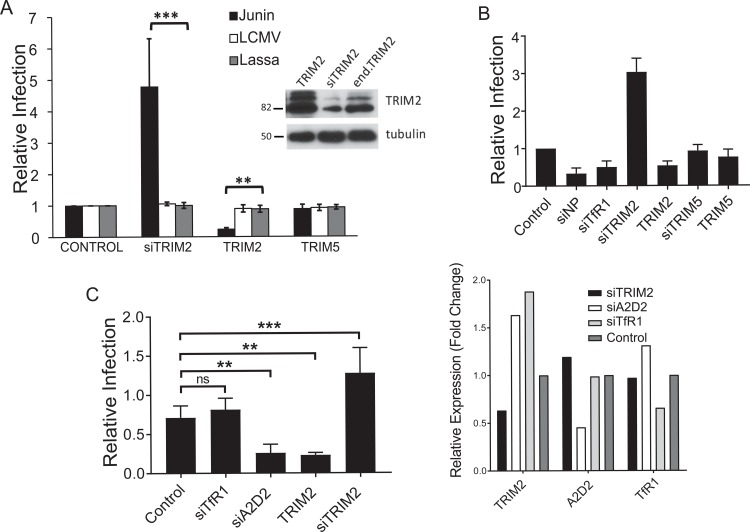
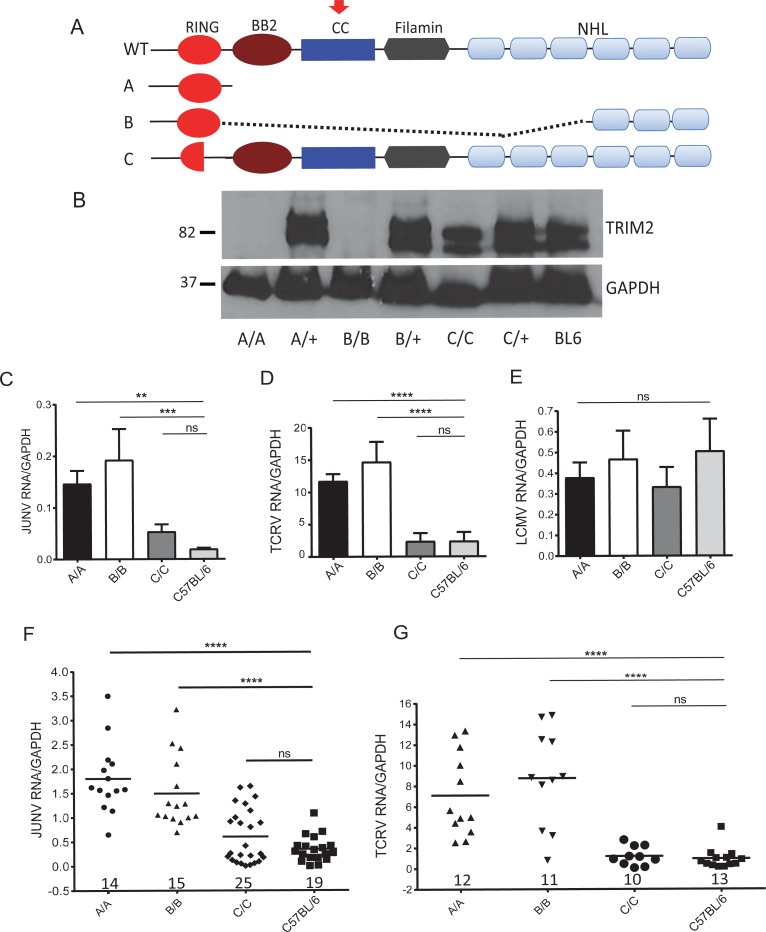
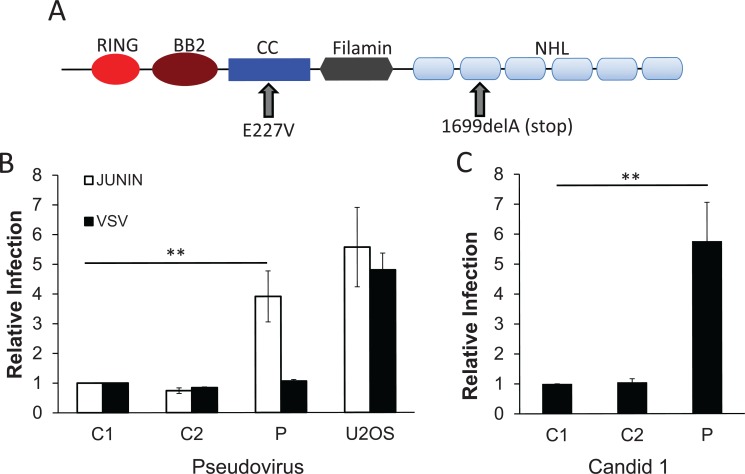
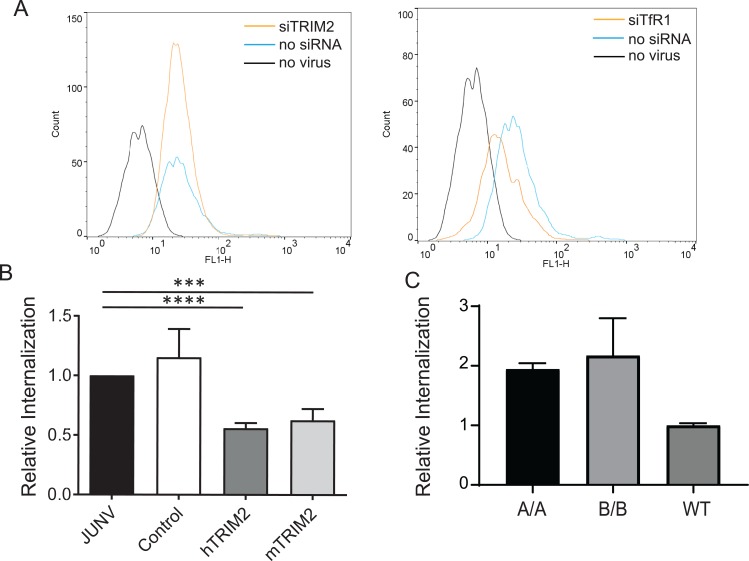
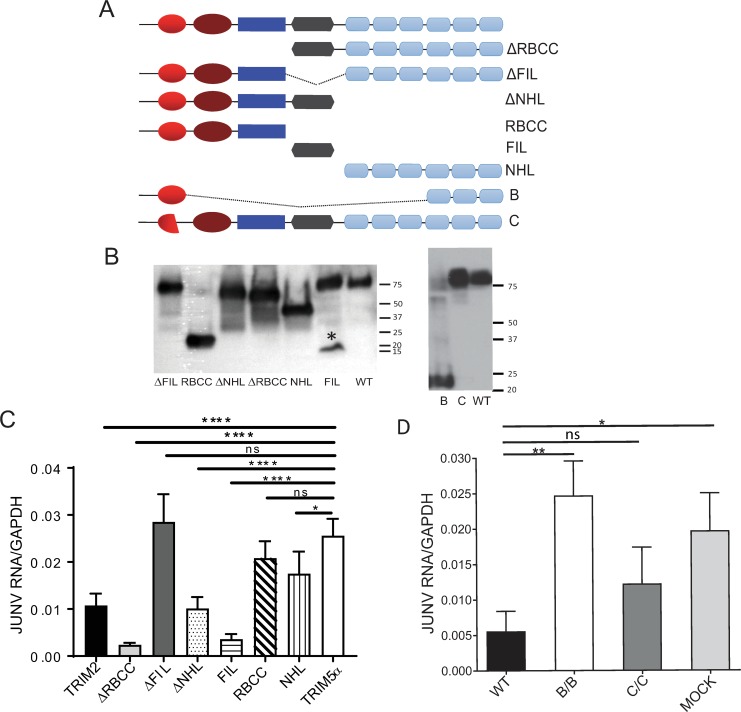
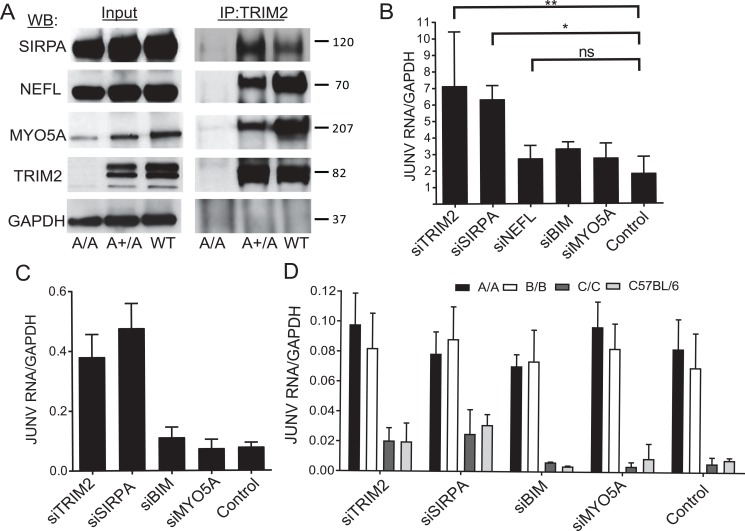
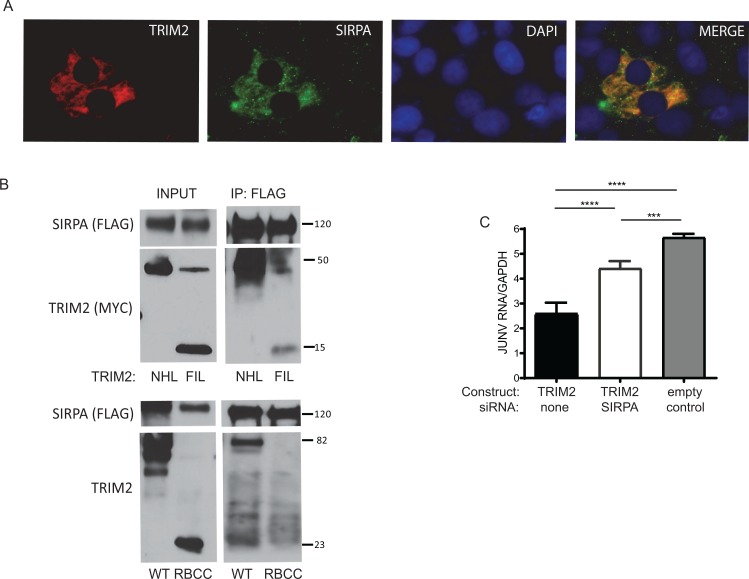
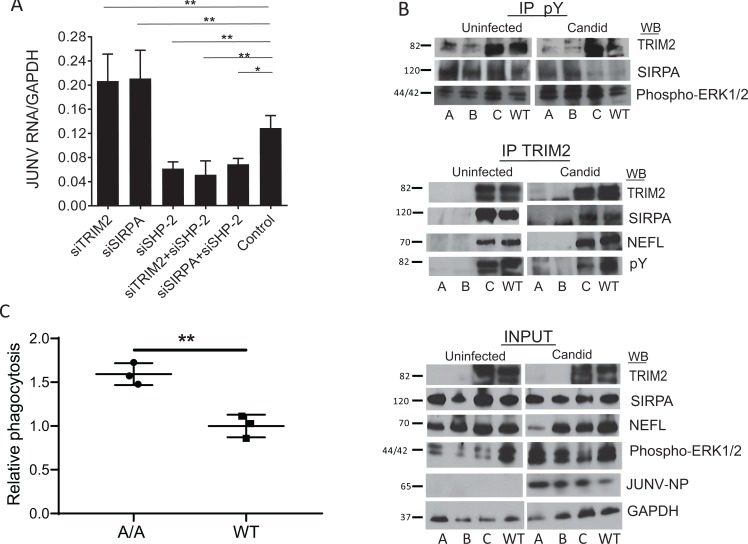
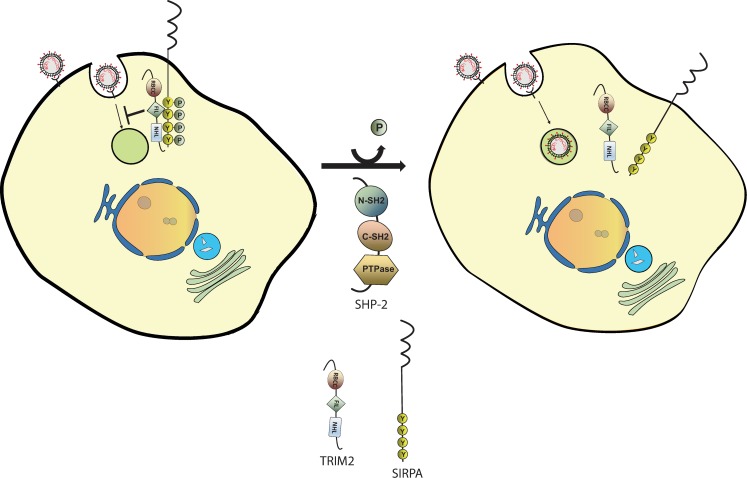
Tripartite motif (TRIM) proteins belong to a large family with many roles in host biology, including restricting virus infection. Here, we found that TRIM2, which has been implicated in cases of Charcot-Marie-Tooth disease (CMTD) in humans, acts by blocking hemorrhagic fever New World arenavirus (NWA) entry into cells. We show that Trim2-knockout mice, as well as primary fibroblasts from a CMTD patient with mutations in TRIM2, are more highly infected by the NWAs Junín and Tacaribe virus than wild-type mice or cells are. Using mice with different Trim2 gene deletions and TRIM2 mutant constructs, we demonstrate that its antiviral activity is uniquely independent of the RING domain encoding ubiquitin ligase activity. Finally, we show that one member of the TRIM2 interactome, signal regulatory protein α (SIRPA), a known inhibitor of phagocytosis, also restricts NWA infection and conversely that TRIM2 limits phagocytosis of apoptotic cells. In addition to demonstrating a novel antiviral mechanism for TRIM proteins, these studies suggest that the NWA entry and phagocytosis pathways overlap.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Identification of a novel Bcl-2-interacting mediator of cell death (Bim) E3 ligase, tripartite motif-containing protein 2 (TRIM2), and its role in rapid ischemic tolerance-induced neuroprotection.J Biol Chem. 2011 Jun 3;286(22):19331-9. doi: 10.1074/jbc.M110.197707. Epub 2011 Apr 8. J Biol Chem. 2011. PMID: 21478148 Free PMC article.
-
The New World arenavirus Tacaribe virus induces caspase-dependent apoptosis in infected cells.J Gen Virol. 2016 Apr;97(4):855-866. doi: 10.1099/jgv.0.000403. Epub 2016 Jan 14. J Gen Virol. 2016. PMID: 26769540
-
Deficiency of the E3 ubiquitin ligase TRIM2 in early-onset axonal neuropathy.Hum Mol Genet. 2013 Aug 1;22(15):2975-83. doi: 10.1093/hmg/ddt149. Epub 2013 Apr 4. Hum Mol Genet. 2013. PMID: 23562820
-
Expression and Role of TRIM2 in Human Diseases.Biomed Res Int. 2022 Aug 23;2022:9430509. doi: 10.1155/2022/9430509. eCollection 2022. Biomed Res Int. 2022. PMID: 36051486 Free PMC article. Review.
-
Pathomechanisms of mutant proteins in Charcot-Marie-Tooth disease.Neuromolecular Med. 2006;8(1-2):217-42. doi: 10.1385/nmm:8:1-2:217. Neuromolecular Med. 2006. PMID: 16775378 Review.
Cited by
-
Ubiquitin Proteasome Gene Signatures in Ependymoma Molecular Subtypes.Int J Mol Sci. 2022 Oct 15;23(20):12330. doi: 10.3390/ijms232012330. Int J Mol Sci. 2022. PMID: 36293188 Free PMC article. Review.
-
The Virus-Host Interplay in Junín Mammarenavirus Infection.Viruses. 2022 May 24;14(6):1134. doi: 10.3390/v14061134. Viruses. 2022. PMID: 35746604 Free PMC article. Review.
-
Neurodevelopmental Disorders (NDD) Caused by Genomic Alterations of the Ubiquitin-Proteasome System (UPS): the Possible Contribution of Immune Dysregulation to Disease Pathogenesis.Front Mol Neurosci. 2021 Sep 8;14:733012. doi: 10.3389/fnmol.2021.733012. eCollection 2021. Front Mol Neurosci. 2021. PMID: 34566579 Free PMC article. Review.
-
The board is set, the pieces are moving: Modulation of New World arenavirus entry by host proteins.PLoS Pathog. 2021 Jun 10;17(6):e1009605. doi: 10.1371/journal.ppat.1009605. eCollection 2021 Jun. PLoS Pathog. 2021. PMID: 34111222 Free PMC article. Review. No abstract available.
-
Hemorrhagic Fever-Causing Arenaviruses: Lethal Pathogens and Potent Immune Suppressors.Front Immunol. 2019 Mar 13;10:372. doi: 10.3389/fimmu.2019.00372. eCollection 2019. Front Immunol. 2019. PMID: 30918506 Free PMC article. Review.
References
-
- Cao W, Henry MD, Borrow P, Yamada H, Elder JH, Ravkov EV, et al. Identification of alpha-dystroglycan as a receptor for lymphocytic choriomeningitis virus and Lassa fever virus. Science. 1998;282(5396):2079–81. Epub 1998/12/16. . - PubMed
-
- Flanagan ML, Oldenburg J, Reignier T, Holt N, Hamilton GA, Martin VK, et al. New World Clade B arenaviruses can use transferrin receptor 1 (TfR1)-dependent and -independent entry pathways, and glycoproteins from human pathogenic strains are associated with the use of TfR1. J Virol. 2008; 82:938–48. 10.1128/JVI.01397-07 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
