

O-GlcNAc Transferase Links Glucose Metabolism to MAVS-Mediated Antiviral Innate Immunity
- PMID: 30543776
- PMCID: PMC6296827
- DOI: 10.1016/j.chom.2018.11.001
O-GlcNAc Transferase Links Glucose Metabolism to MAVS-Mediated Antiviral Innate Immunity
Abstract
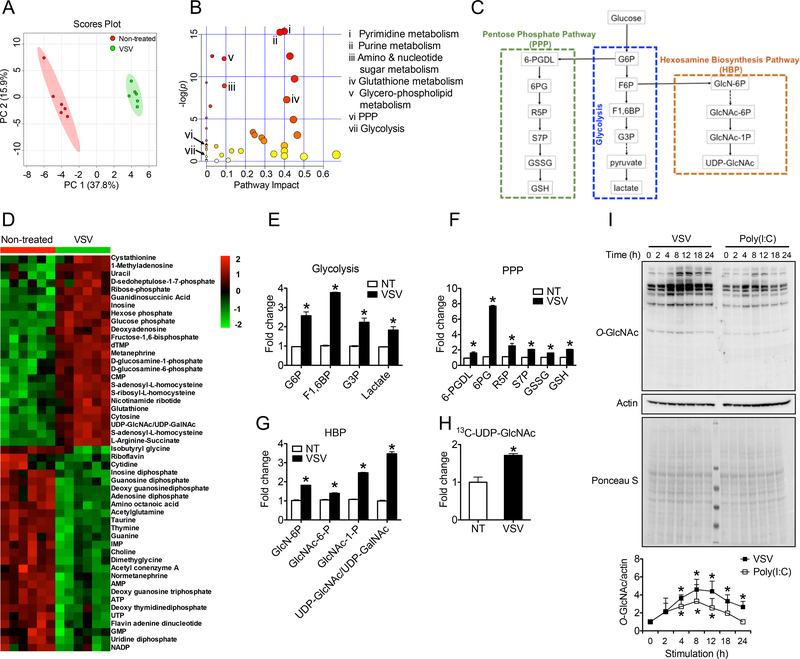
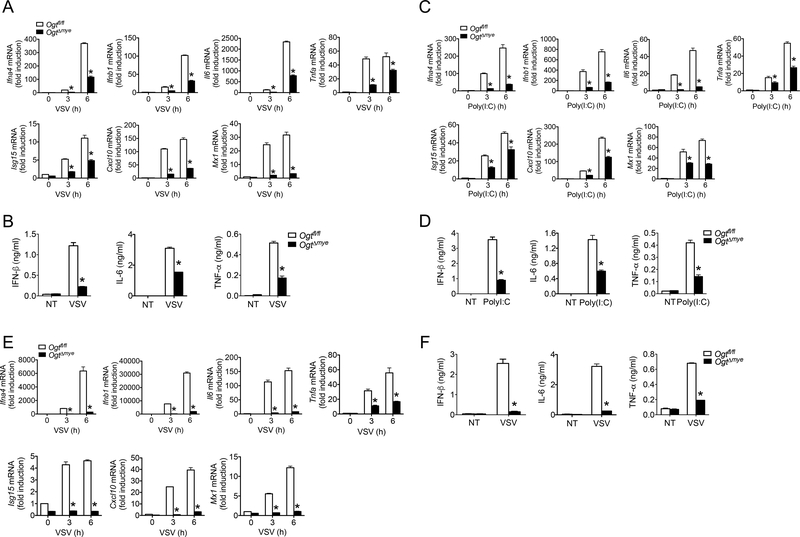
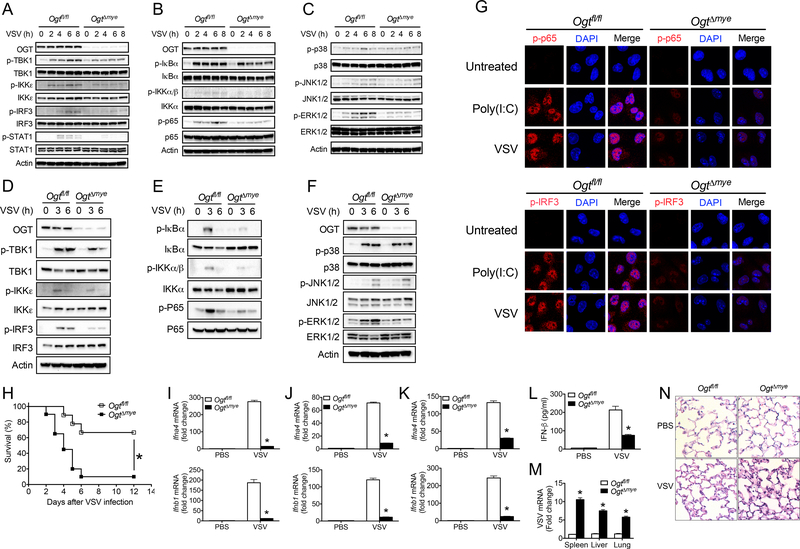
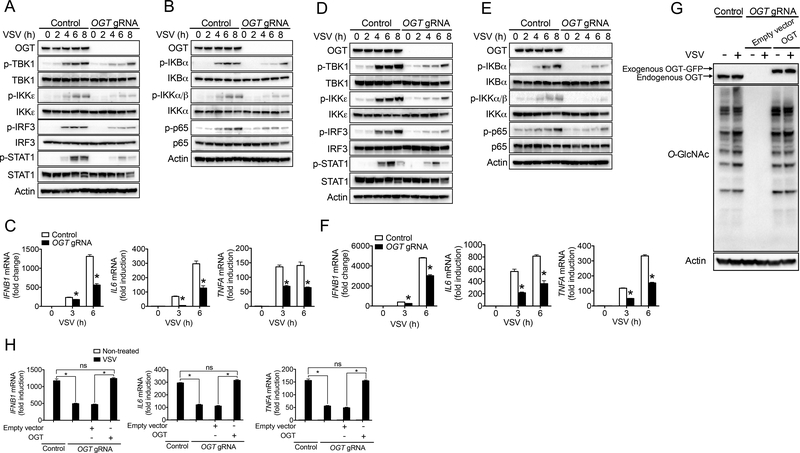
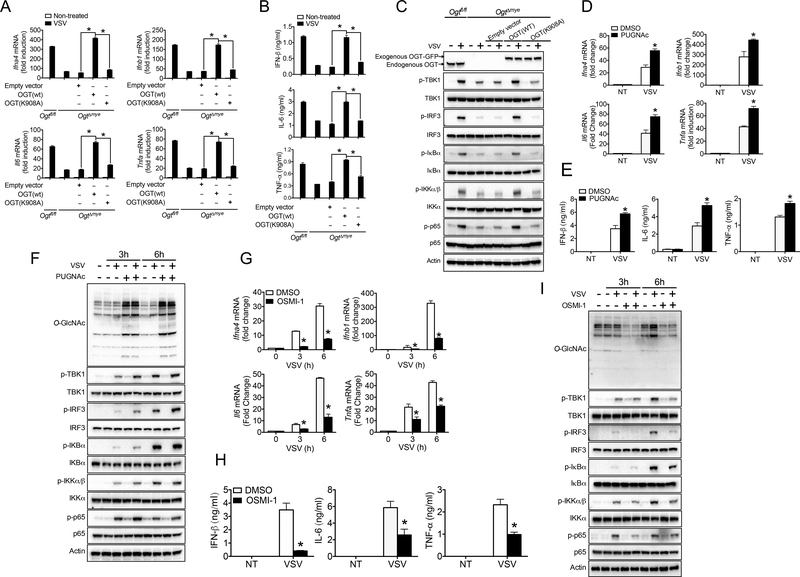
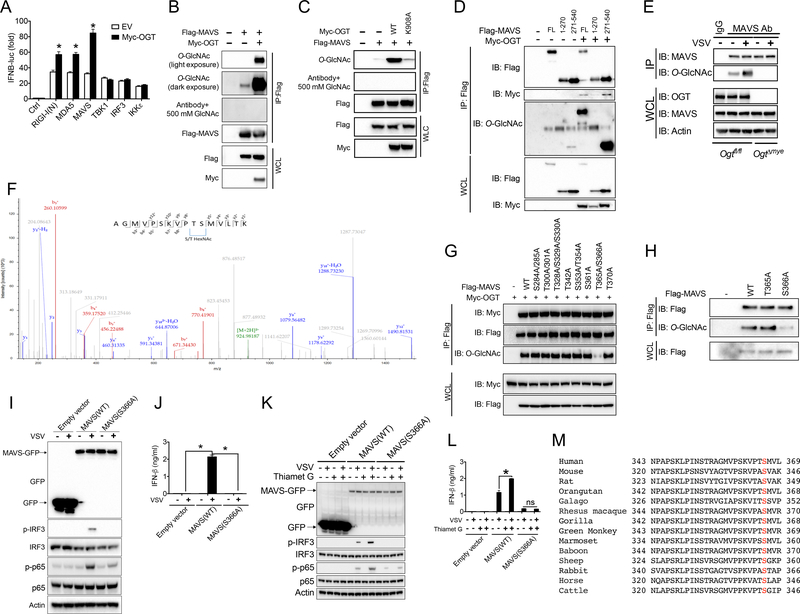
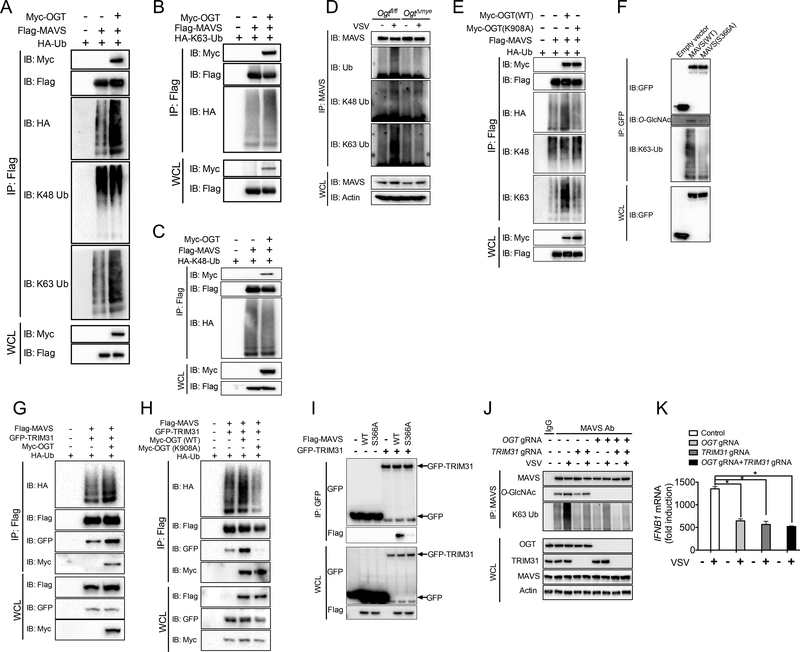
Increased glucose metabolism in immune cells not only serves as a hallmark feature of acute inflammation but also profoundly affects disease outcome following bacterial infection and tissue damage. However, the role of individual glucose metabolic pathways during viral infection remains largely unknown. Here we demonstrate an essential function of the hexosamine biosynthesis pathway (HBP)-associated O-linked β-N-acetylglucosamine (O-GlcNAc) signaling in promoting antiviral innate immunity. Challenge of macrophages with vesicular stomatitis viruses (VSVs) enhances HBP activity and downstream protein O-GlcNAcylation. Human and murine cells deficient of O-GlcNAc transferase, a key enzyme for protein O-GlcNAcylation, show defective antiviral immune responses upon VSV challenge. Mechanistically, O-GlcNAc transferase-mediated O-GlcNAcylation of the signaling adaptor MAVS on serine 366 is required for K63-linked ubiquitination of MAVS and subsequent downstream retinoic-acid inducible gene-like receptor -antiviral signaling activation. Thus, our study identifies a molecular mechanism by which HBP-mediated O-GlcNAcylation regulates MAVS function and highlights the importance of glucose metabolism in antiviral innate immunity.
Keywords: MAVS; O-GlcNAc transferase; antiviral immunity; glucose metabolism; hexosamine biosynthesis pathway (HBP).
Copyright © 2018 Elsevier Inc. All rights reserved.
Conflict of interest statement
DECLARATION OF INTERESTS
The authors declare no competing interests.
Figures
Similar articles
-
Hexosamine biosynthetic pathway promotes the antiviral activity of SAMHD1 by enhancing O-GlcNAc transferase-mediated protein O-GlcNAcylation.Theranostics. 2021 Jan 1;11(2):805-823. doi: 10.7150/thno.50230. eCollection 2021. Theranostics. 2021. PMID: 33391506 Free PMC article.
-
MAVS O-GlcNAcylation Is Essential for Host Antiviral Immunity against Lethal RNA Viruses.Cell Rep. 2019 Aug 27;28(9):2386-2396.e5. doi: 10.1016/j.celrep.2019.07.085. Cell Rep. 2019. PMID: 31461653
-
O-GlcNAc Transferase Suppresses Inflammation and Necroptosis by Targeting Receptor-Interacting Serine/Threonine-Protein Kinase 3.Immunity. 2019 Mar 19;50(3):576-590.e6. doi: 10.1016/j.immuni.2019.01.007. Epub 2019 Feb 12. Immunity. 2019. PMID: 30770249 Free PMC article.
-
Hexosamine biosynthetic pathway and O-GlcNAc cycling of glucose metabolism in brain function and disease.Am J Physiol Cell Physiol. 2023 Oct 1;325(4):C981-C998. doi: 10.1152/ajpcell.00191.2023. Epub 2023 Aug 21. Am J Physiol Cell Physiol. 2023. PMID: 37602414 Review.
-
Protein O-GlcNAcylation Regulates Innate Immune Cell Function.Front Immunol. 2022 Feb 3;13:805018. doi: 10.3389/fimmu.2022.805018. eCollection 2022. Front Immunol. 2022. PMID: 35185892 Free PMC article. Review.
Cited by
-
Virus subtype-specific suppression of MAVS aggregation and activation by PB1-F2 protein of influenza A (H7N9) virus.PLoS Pathog. 2020 Jun 8;16(6):e1008611. doi: 10.1371/journal.ppat.1008611. eCollection 2020 Jun. PLoS Pathog. 2020. PMID: 32511263 Free PMC article.
-
Age-related elevation of O-GlcNAc causes meiotic arrest in male mice.Cell Death Discov. 2023 May 15;9(1):163. doi: 10.1038/s41420-023-01433-x. Cell Death Discov. 2023. PMID: 37188682 Free PMC article.
-
CCL4 participates in the reprogramming of glucose metabolism induced by ALV-J infection in chicken macrophages.Front Microbiol. 2023 Jun 2;14:1205143. doi: 10.3389/fmicb.2023.1205143. eCollection 2023. Front Microbiol. 2023. PMID: 37333648 Free PMC article.
-
MAVS: A Two-Sided CARD Mediating Antiviral Innate Immune Signaling and Regulating Immune Homeostasis.Front Microbiol. 2021 Sep 9;12:744348. doi: 10.3389/fmicb.2021.744348. eCollection 2021. Front Microbiol. 2021. PMID: 34566944 Free PMC article. Review.
-
Regulation of RIG-I-like receptor-mediated signaling: interaction between host and viral factors.Cell Mol Immunol. 2021 Mar;18(3):539-555. doi: 10.1038/s41423-020-00602-7. Epub 2021 Jan 18. Cell Mol Immunol. 2021. PMID: 33462384 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
- R01 CA216853/CA/NCI NIH HHS/United States
- U54 CA163120/CA/NCI NIH HHS/United States
- P30 CA016086/CA/NCI NIH HHS/United States
- P01 CA120964/CA/NCI NIH HHS/United States
- P50 CA127297/CA/NCI NIH HHS/United States
- R01 GM120496/GM/NIGMS NIH HHS/United States
- K99 DE024173/DE/NIDCR NIH HHS/United States
- R01 CA163649/CA/NCI NIH HHS/United States
- R01 CA210439/CA/NCI NIH HHS/United States
- R01 DE026728/DE/NIDCR NIH HHS/United States
- R00 DE024173/DE/NIDCR NIH HHS/United States
- P30 CA006516/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
