

Ubiquitin-dependent switch during assembly of the proteasomal ATPases mediated by Not4 ubiquitin ligase
- PMID: 30530678
- PMCID: PMC6310792
- DOI: 10.1073/pnas.1805353115
Ubiquitin-dependent switch during assembly of the proteasomal ATPases mediated by Not4 ubiquitin ligase
Abstract
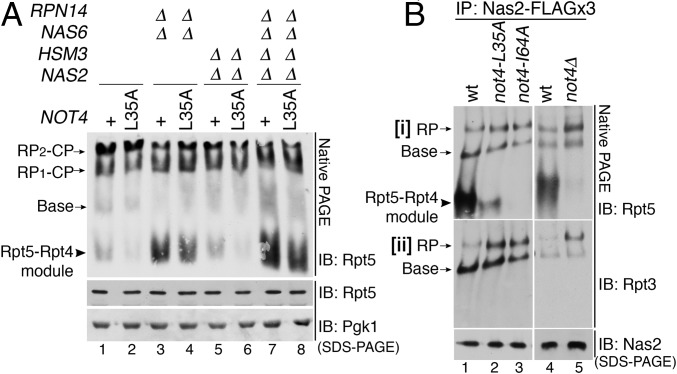
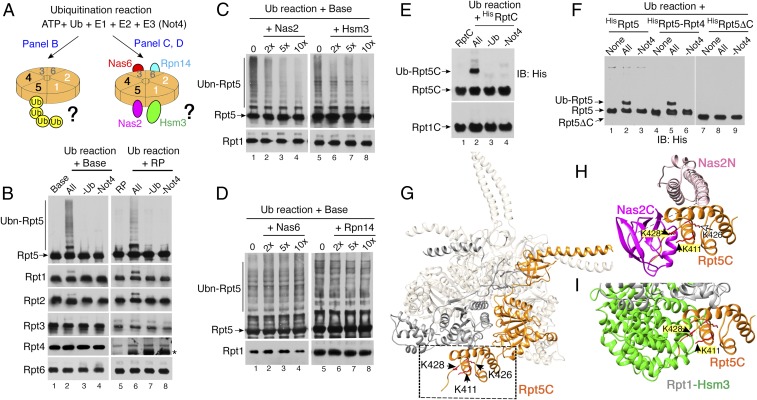
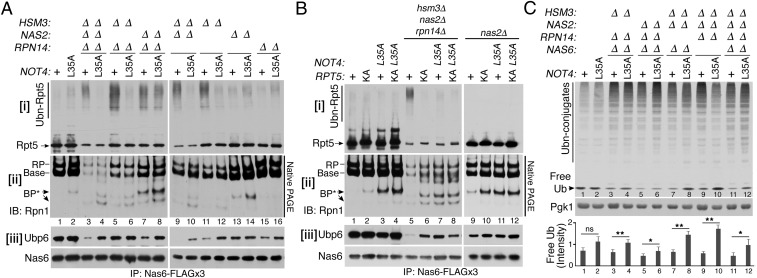
In the proteasome holoenzyme, the hexameric ATPases (Rpt1-Rpt6) enable degradation of ubiquitinated proteins by unfolding and translocating them into the proteolytic core particle. During early-stage proteasome assembly, individual Rpt proteins assemble into the hexameric "Rpt ring" through binding to their cognate chaperones: Nas2, Hsm3, Nas6, and Rpn14. Here, we show that Rpt ring assembly employs a specific ubiquitination-mediated control. An E3 ligase, Not4, selectively ubiquitinates Rpt5 during Rpt ring assembly. To access Rpt5, Not4 competes with Nas2 until the penultimate step and then with Hsm3 at the final step of Rpt ring completion. Using the known Rpt-chaperone cocrystal structures, we show that Not4-mediated ubiquitination sites in Rpt5 are obstructed by Nas2 and Hsm3. Thus, Not4 can distinguish a Rpt ring that matures without these chaperones, based on its accessibility to Rpt5. Rpt5 ubiquitination does not destabilize the ring but hinders incorporation of incoming subunits-Rpn1 ubiquitin receptor and Ubp6 deubiquitinase-thereby blocking progression of proteasome assembly and ubiquitin regeneration from proteasome substrates. Our findings reveal an assembly checkpoint where Not4 monitors chaperone actions during hexameric ATPase ring assembly, thereby ensuring the accuracy of proteasome holoenzyme maturation.
Keywords: AAA+ ATPase; Not4; assembly chaperone; checkpoint; proteasome.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

Similar articles
-
Two alternative mechanisms regulate the onset of chaperone-mediated assembly of the proteasomal ATPases.J Biol Chem. 2019 Apr 19;294(16):6562-6577. doi: 10.1074/jbc.RA118.006298. Epub 2019 Feb 27. J Biol Chem. 2019. PMID: 30814255 Free PMC article.
-
Reconfiguration of the proteasome during chaperone-mediated assembly.Nature. 2013 May 23;497(7450):512-6. doi: 10.1038/nature12123. Epub 2013 May 5. Nature. 2013. PMID: 23644457 Free PMC article.
-
Proteasome Activation is Mediated via a Functional Switch of the Rpt6 C-terminal Tail Following Chaperone-dependent Assembly.Sci Rep. 2015 Oct 9;5:14909. doi: 10.1038/srep14909. Sci Rep. 2015. PMID: 26449534 Free PMC article.
-
Assembly manual for the proteasome regulatory particle: the first draft.Biochem Soc Trans. 2010 Feb;38(Pt 1):6-13. doi: 10.1042/BST0380006. Biochem Soc Trans. 2010. PMID: 20074027 Free PMC article. Review.
-
Ubiquitin recognition by the proteasome.J Biochem. 2017 Feb 1;161(2):113-124. doi: 10.1093/jb/mvw091. J Biochem. 2017. PMID: 28069863 Review.
Cited by
-
Conserved proline residues in the coiled coil-OB domain linkers of Rpt proteins facilitate eukaryotic proteasome base assembly.J Biol Chem. 2021 Jan-Jun;296:100660. doi: 10.1016/j.jbc.2021.100660. Epub 2021 Apr 14. J Biol Chem. 2021. PMID: 33862083 Free PMC article.
-
The Ccr4-Not complex regulates TORC1 signaling and mitochondrial metabolism by promoting vacuole V-ATPase activity.PLoS Genet. 2020 Oct 16;16(10):e1009046. doi: 10.1371/journal.pgen.1009046. eCollection 2020 Oct. PLoS Genet. 2020. PMID: 33064727 Free PMC article.
-
Ccr4-Not as a mediator of environmental signaling: a jack of all trades and master of all.Curr Genet. 2021 Oct;67(5):707-713. doi: 10.1007/s00294-021-01180-5. Epub 2021 Mar 31. Curr Genet. 2021. PMID: 33791857 Free PMC article. Review.
-
Assembly checkpoint of the proteasome regulatory particle is activated by coordinated actions of proteasomal ATPase chaperones.Cell Rep. 2022 Jun 7;39(10):110918. doi: 10.1016/j.celrep.2022.110918. Cell Rep. 2022. PMID: 35675778 Free PMC article.
-
Dynamic Regulation of the 26S Proteasome: From Synthesis to Degradation.Front Mol Biosci. 2019 Jun 7;6:40. doi: 10.3389/fmolb.2019.00040. eCollection 2019. Front Mol Biosci. 2019. PMID: 31231659 Free PMC article. Review.
References
-
- da Fonseca PC, He J, Morris EP. Molecular model of the human 26S proteasome. Mol Cell. 2012;46:54–66. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
