

The Viral Tegument Protein pp65 Impairs Transcriptional Upregulation of IL-1β by Human Cytomegalovirus through Inhibition of NF-kB Activity
- PMID: 30332797
- PMCID: PMC6213739
- DOI: 10.3390/v10100567
The Viral Tegument Protein pp65 Impairs Transcriptional Upregulation of IL-1β by Human Cytomegalovirus through Inhibition of NF-kB Activity
Abstract
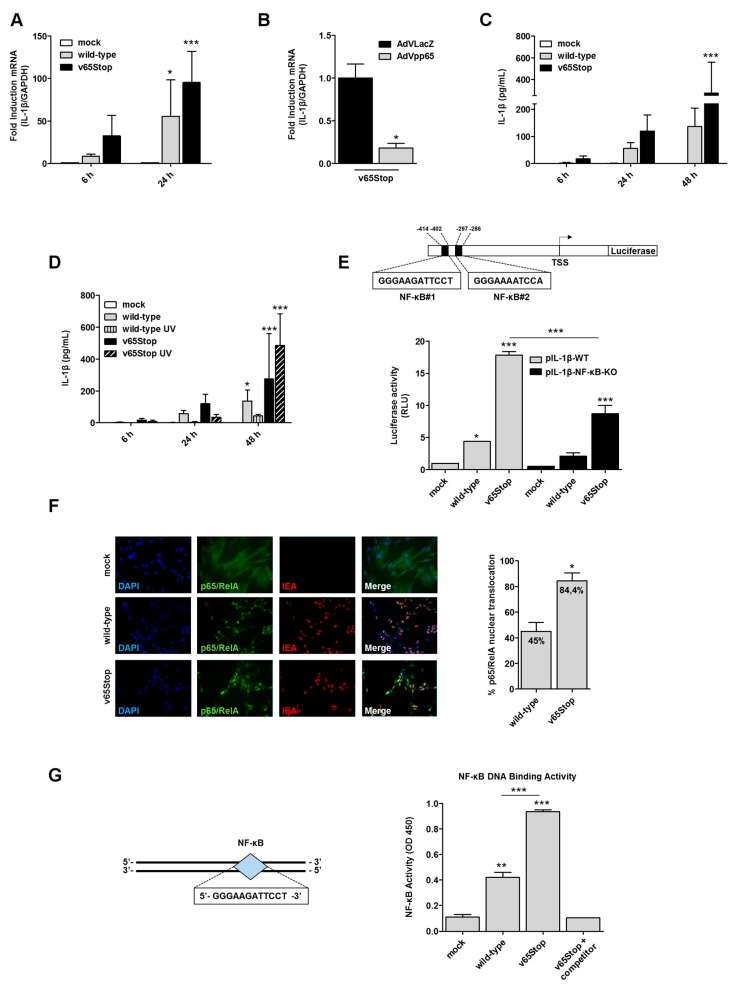
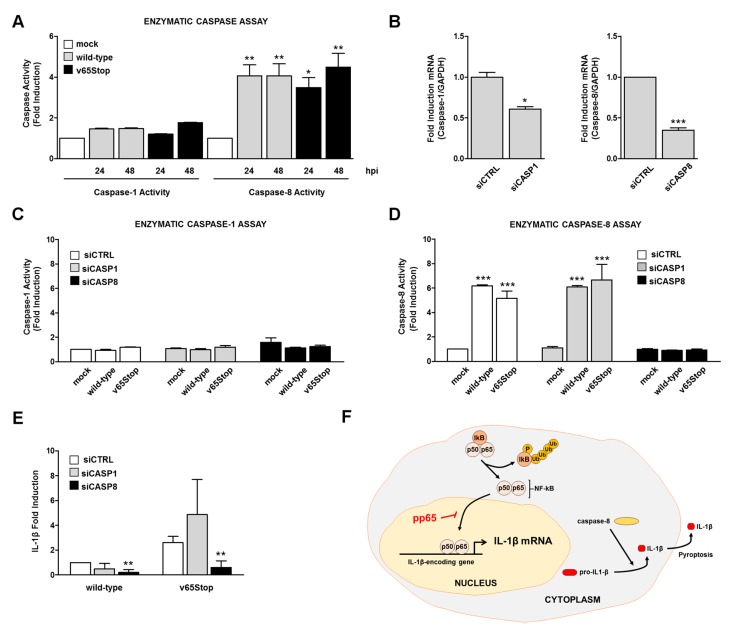
Interleukin-1β (IL-1β) is a key effector of the inflammasome complex in response to pathogens and danger signals. Although it is well known that assembly of the inflammasome triggers proteolytic cleavage of the biologically inactive precursor pro-IL-1β into its mature secreted form, the mechanism by which human cytomegalovirus (HCMV) regulates IL-1β production via the inflammasome is still poorly understood. Here, we show that the infection of human foreskin fibroblasts (HFFs) with a mutant HCMV lacking the tegument protein pp65 (v65Stop) results in higher expression levels of mature IL-1β compared to its wild-type counterpart, suggesting that pp65 mediates HCMV immune evasion through downmodulation of IL-1β. Furthermore, we show that enhanced IL-1β production by the v65Stop mutant is due in part to induction of DNA binding and the transcriptional activity of NF-κB. Lastly, we demonstrate that HCMV infection of HFFs triggers a non-canonical IL-1β activation pathway where caspase-8 promotes IL-1β maturation independently of caspase-1. Altogether, our findings provide novel mechanistic insights into the interplay between HCMV and the inflammasome system and raise the possibility of targeting pp65 to treat HCMV infection.
Keywords: caspase-8; human cytomegalovirus (HCMV); inflammasome; interleukin-1β (IL-1β); pp65.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Human Cytomegalovirus Tegument Protein pp65 (pUL83) Dampens Type I Interferon Production by Inactivating the DNA Sensor cGAS without Affecting STING.J Virol. 2018 Feb 26;92(6):e01774-17. doi: 10.1128/JVI.01774-17. Print 2018 Mar 15. J Virol. 2018. PMID: 29263269 Free PMC article.
-
Interaction between HCMV pUL83 and human AIM2 disrupts the activation of the AIM2 inflammasome.Virol J. 2017 Feb 20;14(1):34. doi: 10.1186/s12985-016-0673-5. Virol J. 2017. PMID: 28219398 Free PMC article.
-
Regulatory Interaction between the Cellular Restriction Factor IFI16 and Viral pp65 (pUL83) Modulates Viral Gene Expression and IFI16 Protein Stability.J Virol. 2016 Aug 26;90(18):8238-50. doi: 10.1128/JVI.00923-16. Print 2016 Sep 15. J Virol. 2016. PMID: 27384655 Free PMC article.
-
The human cytomegalovirus tegument protein pp65 (pUL83): a key player in innate immune evasion.New Microbiol. 2018 Apr;41(2):87-94. Epub 2018 Jan 31. New Microbiol. 2018. PMID: 29384558 Review.
-
Multifaceted evasion of the interferon response by cytomegalovirus.J Interferon Cytokine Res. 2009 Sep;29(9):609-19. doi: 10.1089/jir.2009.0064. J Interferon Cytokine Res. 2009. PMID: 19708810 Free PMC article. Review.
Cited by
-
Human cytomegalovirus infection triggers a paracrine senescence loop in renal epithelial cells.Commun Biol. 2024 Mar 8;7(1):292. doi: 10.1038/s42003-024-05957-5. Commun Biol. 2024. PMID: 38459109 Free PMC article.
-
Human Cytomegalovirus Primary Infection and Reactivation: Insights From Virion-Carried Molecules.Front Microbiol. 2020 Jul 14;11:1511. doi: 10.3389/fmicb.2020.01511. eCollection 2020. Front Microbiol. 2020. PMID: 32765441 Free PMC article. Review.
-
Emerging Role of PYHIN Proteins as Antiviral Restriction Factors.Viruses. 2020 Dec 18;12(12):1464. doi: 10.3390/v12121464. Viruses. 2020. PMID: 33353088 Free PMC article. Review.
-
The battle between host antiviral innate immunity and immune evasion by cytomegalovirus.Cell Mol Life Sci. 2024 Aug 9;81(1):341. doi: 10.1007/s00018-024-05369-y. Cell Mol Life Sci. 2024. PMID: 39120730 Free PMC article. Review.
-
Insight for Immunotherapy of HCMV Infection.Int J Biol Sci. 2021 Jul 13;17(11):2899-2911. doi: 10.7150/ijbs.58127. eCollection 2021. Int J Biol Sci. 2021. PMID: 34345215 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
- 2015W729WH/Ministero dell'Istruzione, dell'Università e della Ricerca/International
- 2015RMNSTA/Ministero dell'Istruzione, dell'Università e della Ricerca/International
- CSP 2016/Compagnia di San Paolo/International
- LANSRILO1701/Università degli Studi di Torino/International
- DELVRILO1701/Università degli Studi di Torino/International
