

Arabidopsis RNA processing factor SERRATE regulates the transcription of intronless genes
- PMID: 30152752
- PMCID: PMC6135607
- DOI: 10.7554/eLife.37078
Arabidopsis RNA processing factor SERRATE regulates the transcription of intronless genes
Abstract
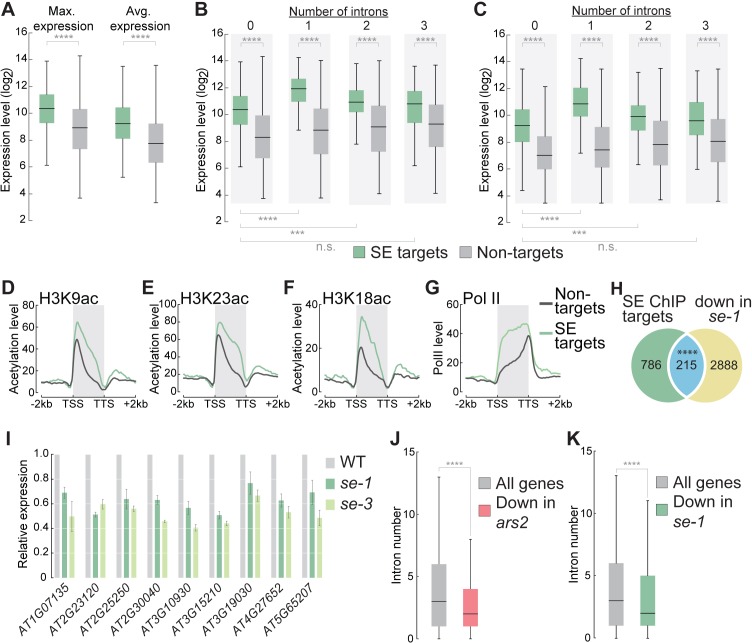
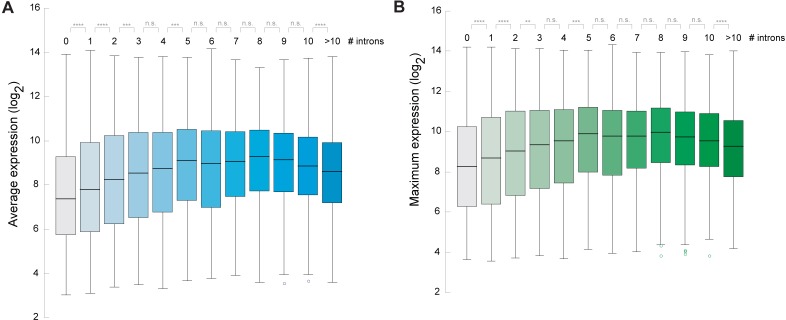
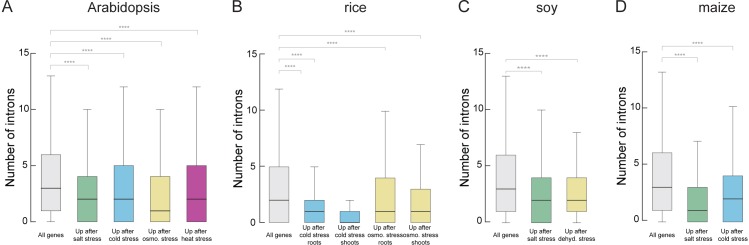
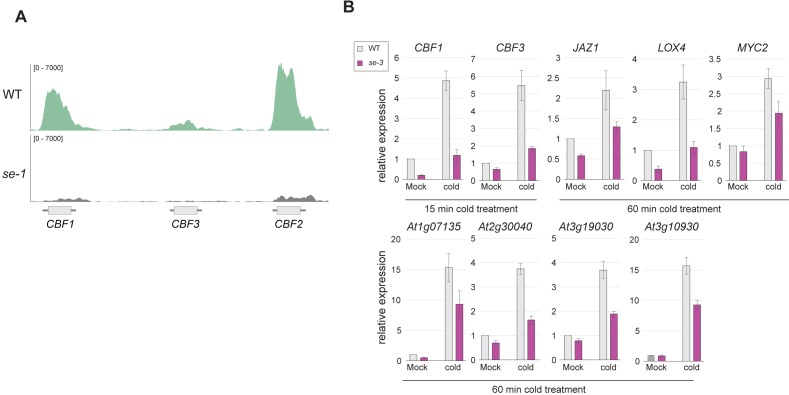
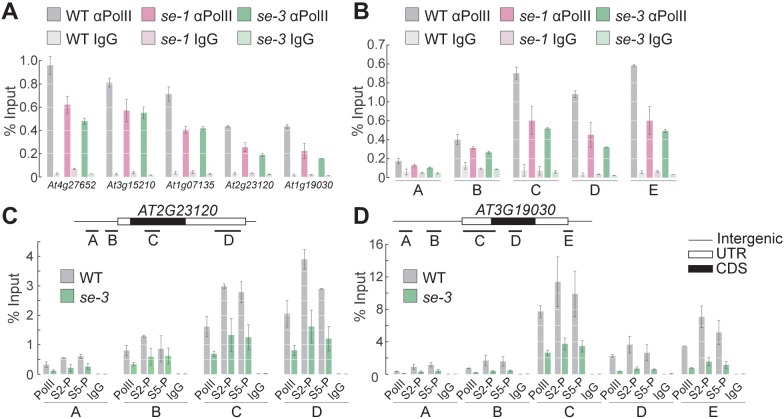
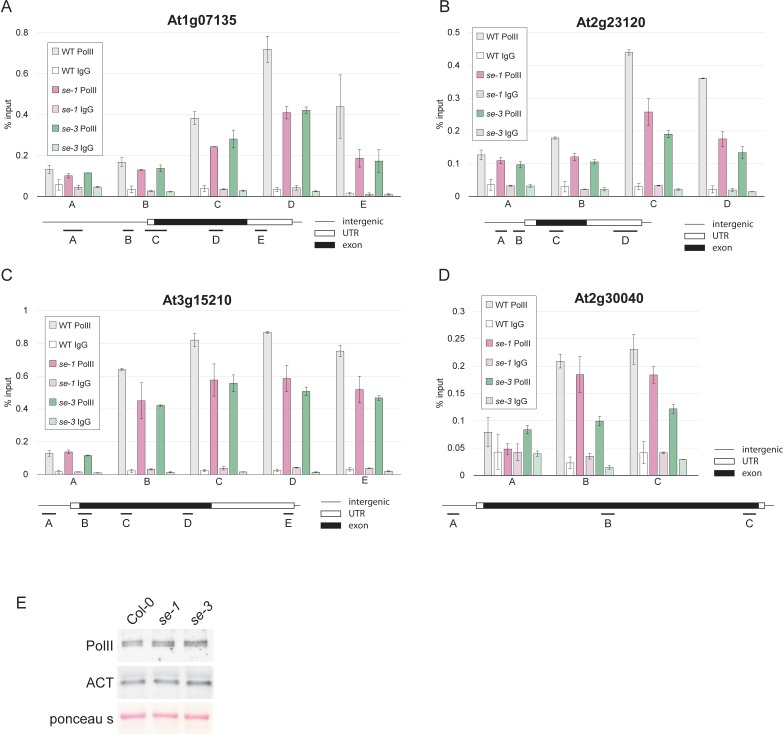
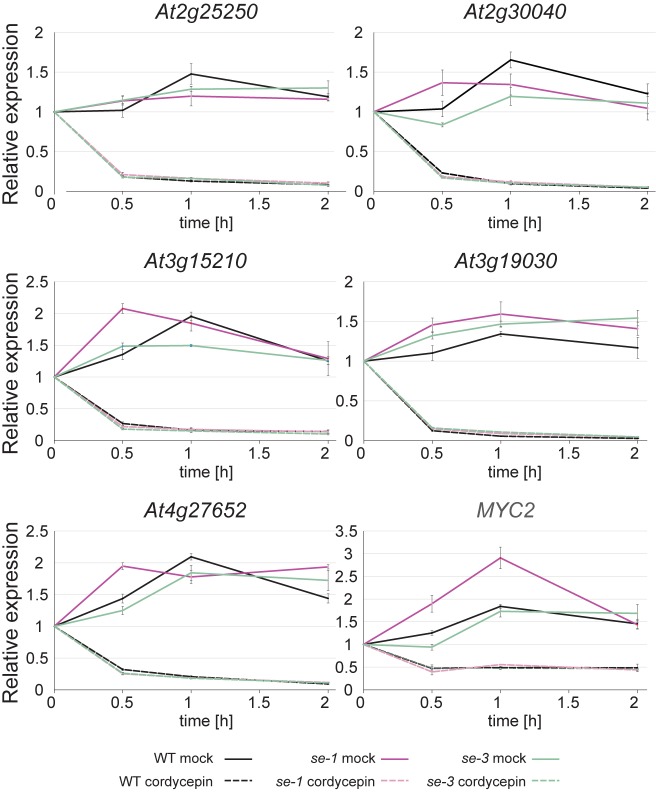
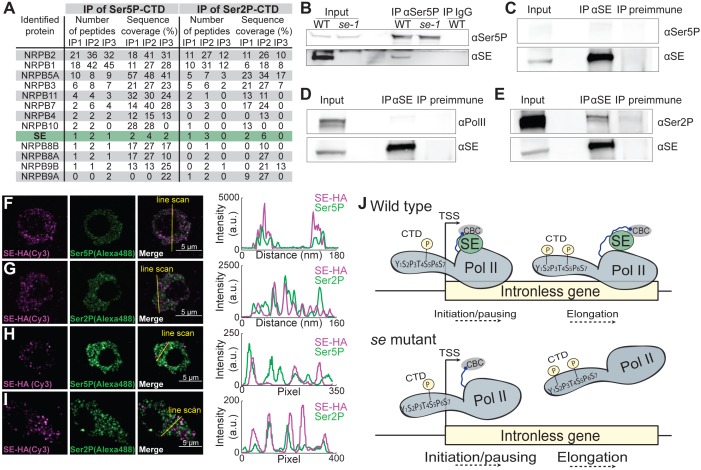
Intron splicing increases proteome complexity, promotes RNA stability, and enhances transcription. However, introns and the concomitant need for splicing extend the time required for gene expression and can cause an undesirable delay in the activation of genes. Here, we show that the plant microRNA processing factor SERRATE (SE) plays an unexpected and pivotal role in the regulation of intronless genes. Arabidopsis SE associated with more than 1000, mainly intronless, genes in a transcription-dependent manner. Chromatin-bound SE liaised with paused and elongating polymerase II complexes and promoted their association with intronless target genes. Our results indicate that stress-responsive genes contain no or few introns, which negatively affects their expression strength, but that some genes circumvent this limitation via a novel SE-dependent transcriptional activation mechanism. Transcriptome analysis of a Drosophila mutant defective in ARS2, the metazoan homologue of SE, suggests that SE/ARS2 function in regulating intronless genes might be conserved across kingdoms.
Keywords: A. thaliana; D. melanogaster; SERRATE; chromosomes; gene expression; intron; intronless genes; plant biology; splicing; transcription.
© 2018, Speth et al.
Conflict of interest statement
CS, ES, CM, SC, Sz, SR, ID, BM, YS, MS, CL, SL No competing interests declared
Figures




Similar articles
-
Intron splicing suppresses RNA silencing in Arabidopsis.Plant J. 2011 Oct;68(1):159-67. doi: 10.1111/j.1365-313X.2011.04676.x. Epub 2011 Aug 5. Plant J. 2011. PMID: 21689169
-
Transcriptome-Wide Identification of RNA Targets of Arabidopsis SERINE/ARGININE-RICH45 Uncovers the Unexpected Roles of This RNA Binding Protein in RNA Processing.Plant Cell. 2015 Dec;27(12):3294-308. doi: 10.1105/tpc.15.00641. Epub 2015 Nov 24. Plant Cell. 2015. PMID: 26603559 Free PMC article.
-
CDKF;1 and CDKD protein kinases regulate phosphorylation of serine residues in the C-terminal domain of Arabidopsis RNA polymerase II.Plant Cell. 2012 Apr;24(4):1626-42. doi: 10.1105/tpc.112.096834. Epub 2012 Apr 30. Plant Cell. 2012. PMID: 22547781 Free PMC article.
-
SERRATE: a key factor in coordinated RNA processing in plants.Trends Plant Sci. 2023 Jul;28(7):841-853. doi: 10.1016/j.tplants.2023.03.009. Epub 2023 Apr 4. Trends Plant Sci. 2023. PMID: 37019716 Review.
-
Regulated RNA processing in the control of Arabidopsis flowering.Int J Dev Biol. 2005;49(5-6):773-80. doi: 10.1387/ijdb.051995vq. Int J Dev Biol. 2005. PMID: 16096981 Review.
Cited by
-
Chromatin-associated microprocessor assembly is regulated by the U1 snRNP auxiliary protein PRP40.Plant Cell. 2022 Nov 29;34(12):4920-4935. doi: 10.1093/plcell/koac278. Plant Cell. 2022. PMID: 36087009 Free PMC article.
-
The Arabidopsis U1 snRNP regulates mRNA 3'-end processing.Nat Plants. 2024 Oct;10(10):1514-1531. doi: 10.1038/s41477-024-01796-8. Epub 2024 Sep 23. Nat Plants. 2024. PMID: 39313562 Free PMC article.
-
The Intrinsically Disordered Protein CARP9 Bridges HYL1 to AGO1 in the Nucleus to Promote MicroRNA Activity.Plant Physiol. 2020 Sep;184(1):316-329. doi: 10.1104/pp.20.00258. Epub 2020 Jul 7. Plant Physiol. 2020. PMID: 32636339 Free PMC article.
-
Capture of regulatory factors via CRISPR-dCas9 for mechanistic analysis of fine-tuned SERRATE expression in Arabidopsis.Nat Plants. 2024 Jan;10(1):86-99. doi: 10.1038/s41477-023-01575-x. Epub 2024 Jan 2. Nat Plants. 2024. PMID: 38168608
-
MAC5, an RNA-binding protein, protects pri-miRNAs from SERRATE-dependent exoribonuclease activities.Proc Natl Acad Sci U S A. 2020 Sep 22;117(38):23982-23990. doi: 10.1073/pnas.2008283117. Epub 2020 Sep 4. Proc Natl Acad Sci U S A. 2020. PMID: 32887800 Free PMC article.
References
-
- Ahlmann-Eltze C. significance brackets for ggplot2. Ggsignif 2017
-
- Belamkar V, Weeks NT, Bharti AK, Farmer AD, Graham MA, Cannon SB. Comprehensive characterization and RNA-Seq profiling of the HD-Zip transcription factor family in soybean (Glycine max) during dehydration and salt stress. BMC Genomics. 2014;15:950. doi: 10.1186/1471-2164-15-950. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
