

Murine CMV Expressing the High Affinity NKG2D Ligand MULT-1: A Model for the Development of Cytomegalovirus-Based Vaccines
- PMID: 29867968
- PMCID: PMC5949336
- DOI: 10.3389/fimmu.2018.00991
Murine CMV Expressing the High Affinity NKG2D Ligand MULT-1: A Model for the Development of Cytomegalovirus-Based Vaccines
Abstract
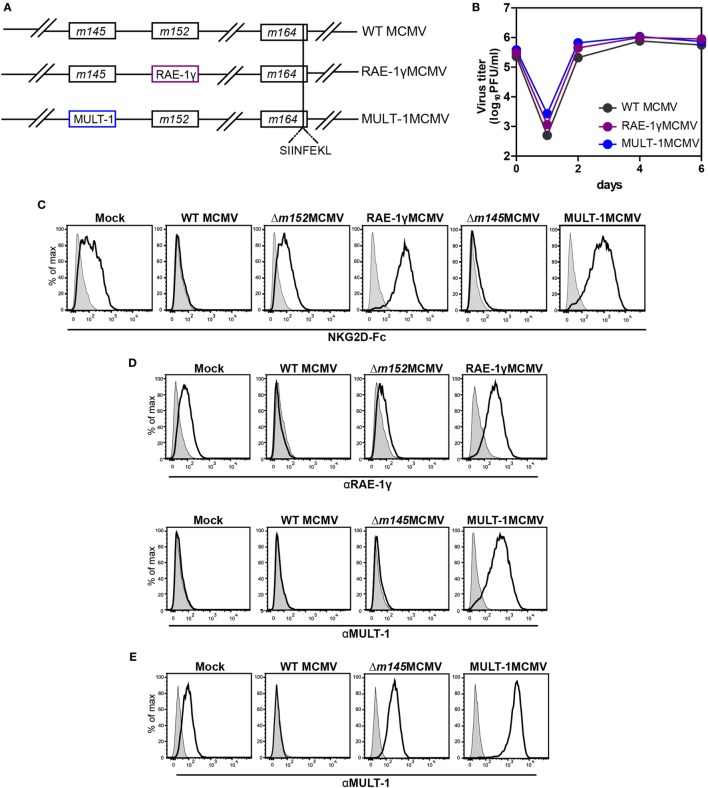
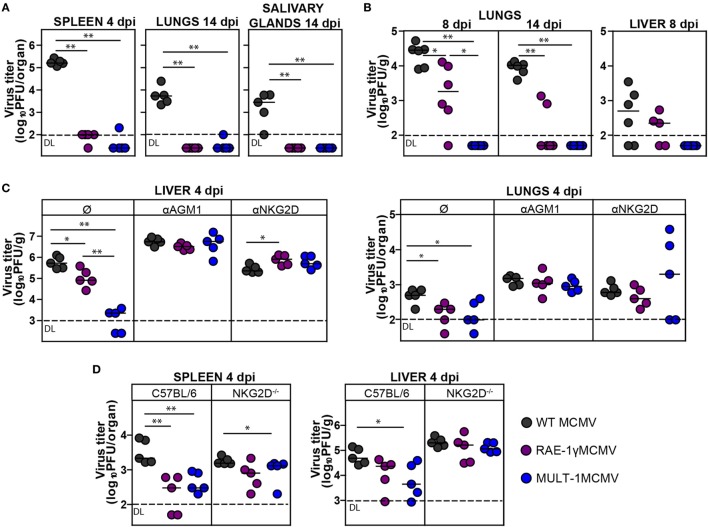
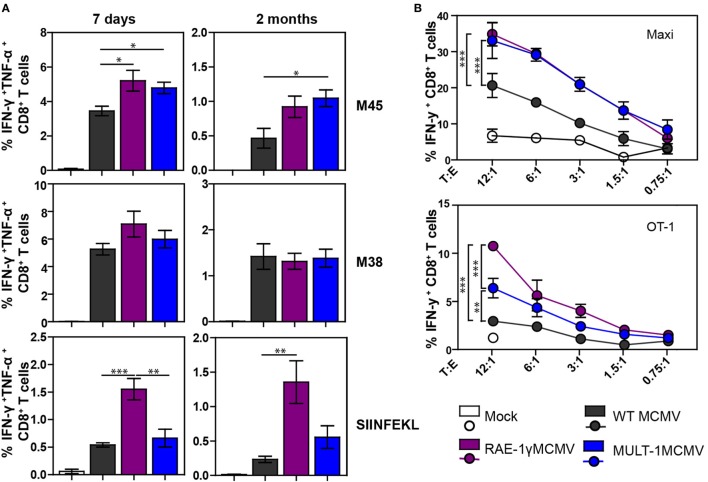
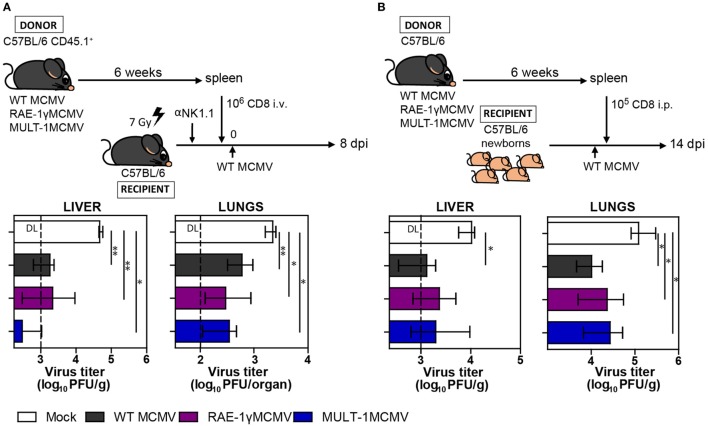
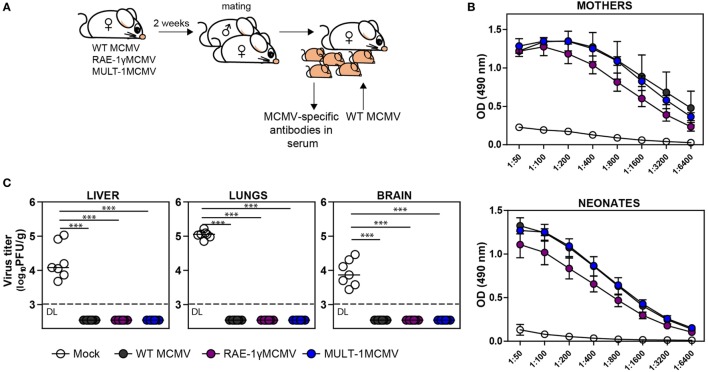
The development of a vaccine against human cytomegalovirus (CMV) has been a subject of long-term medical interest. The research during recent years identified CMV as an attractive vaccine vector against infectious diseases and tumors. The immune response to CMV persists over a lifetime and its unique feature is the inflationary T cell response to certain viral epitopes. CMV encodes numerous genes involved in immunoevasion, which are non-essential for virus growth in vitro. The deletion of those genes results in virus attenuation in vivo, which enables us to dramatically manipulate its virulence and the immune response. We have previously shown that the murine CMV (MCMV) expressing RAE-1γ, one of the cellular ligands for the NKG2D receptor, is highly attenuated in vivo but retains the ability to induce a strong CD8+ T cell response. Here, we demonstrate that recombinant MCMV expressing high affinity NKG2D ligand murine UL16 binding protein-like transcript (MULT-1) (MULT-1MCMV) inserted in the place of its viral inhibitor is dramatically attenuated in vivo in a NK cell-dependent manner, both in immunocompetent adult mice and in immunologically immature newborns. MULT-1MCMV was more attenuated than the recombinant virus expressing RAE-1γ. Despite the drastic sensitivity to innate immune control, MULT-1MCMV induced an efficient CD8+ T cell response to viral and vectored antigens. By using in vitro assay, we showed that similar to RAE-1γMCMV, MULT-1 expressing virus provided strong priming of CD8+ T cells. Moreover, MULT-1MCMV was able to induce anti-viral antibodies, which after passing the transplacental barrier protect offspring of immunized mothers from challenge infection. Altogether, this study further supports the concept that CMV expressing NKG2D ligand possesses excellent characteristics to serve as a vaccine or vaccine vector.
Keywords: CD8+ T cells; NK cells; NKG2D; cytomegalovirus; murine CMV; murine UL16 binding protein-like transcript 1; vaccine.
Figures
Similar articles
-
Memory CD8 T Cells Generated by Cytomegalovirus Vaccine Vector Expressing NKG2D Ligand Have Effector-Like Phenotype and Distinct Functional Features.Front Immunol. 2021 Jun 3;12:681380. doi: 10.3389/fimmu.2021.681380. eCollection 2021. Front Immunol. 2021. PMID: 34168650 Free PMC article.
-
Recombinant mouse cytomegalovirus expressing a ligand for the NKG2D receptor is attenuated and has improved vaccine properties.J Clin Invest. 2010 Dec;120(12):4532-45. doi: 10.1172/JCI43961. Epub 2010 Nov 22. J Clin Invest. 2010. PMID: 21099111 Free PMC article.
-
Superior induction and maintenance of protective CD8 T cells in mice infected with mouse cytomegalovirus vector expressing RAE-1γ.Proc Natl Acad Sci U S A. 2013 Oct 8;110(41):16550-5. doi: 10.1073/pnas.1310215110. Epub 2013 Sep 19. Proc Natl Acad Sci U S A. 2013. PMID: 24052528 Free PMC article.
-
Murine cytomegalovirus regulation of NKG2D ligands.Med Microbiol Immunol. 2008 Jun;197(2):159-66. doi: 10.1007/s00430-008-0080-7. Epub 2008 Feb 8. Med Microbiol Immunol. 2008. PMID: 18259774 Review.
-
MHC class I immune evasion in MCMV infection.Med Microbiol Immunol. 2008 Jun;197(2):191-204. doi: 10.1007/s00430-008-0089-y. Epub 2008 Mar 11. Med Microbiol Immunol. 2008. PMID: 18330598 Review.
Cited by
-
Vaccine vectors: the bright side of cytomegalovirus.Med Microbiol Immunol. 2019 Aug;208(3-4):349-363. doi: 10.1007/s00430-019-00597-7. Epub 2019 Mar 21. Med Microbiol Immunol. 2019. PMID: 30900089 Review.
-
Development and Applications of Viral Vectored Vaccines to Combat Zoonotic and Emerging Public Health Threats.Vaccines (Basel). 2020 Nov 13;8(4):680. doi: 10.3390/vaccines8040680. Vaccines (Basel). 2020. PMID: 33202961 Free PMC article. Review.
-
Role of antibodies in confining cytomegalovirus after reactivation from latency: three decades' résumé.Med Microbiol Immunol. 2019 Aug;208(3-4):415-429. doi: 10.1007/s00430-019-00600-1. Epub 2019 Mar 28. Med Microbiol Immunol. 2019. PMID: 30923898 Free PMC article. Review.
-
Fuel and brake of memory T cell inflation.Med Microbiol Immunol. 2019 Aug;208(3-4):329-338. doi: 10.1007/s00430-019-00587-9. Epub 2019 Mar 9. Med Microbiol Immunol. 2019. PMID: 30852648 Review.
-
Cytomegalovirus Infection and Inflammation in Developing Brain.Viruses. 2021 Jun 4;13(6):1078. doi: 10.3390/v13061078. Viruses. 2021. PMID: 34200083 Free PMC article. Review.
References
-
- Boppana SB, Fowler KB. Chapter 36: Persistence in the population: epidemiology and transmission. In: Arvin A, Campadelli-Fiume G, Mocarski E, Moore PS, Roizman B, Whitley R, Yamanishi K, editors. Human Herpesviruses: Biology, Therapy, and Immunoprophylaxis. Cambridge: Cambridge University Press; (2007). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials